Authors and Reviewers
Authors and Reviewers

Table
1.
Discovery of cardiac natriuretic peptides and the heart-specific miR-208a/MED13 axis
| Molecules investigated; Research question; Authors | Author's main results and conclusions |
| ANP; What are the acute renal effects of the extract of rat atrial myocardium? (de Bold AJ et al., 1981)[2] | The atrial extract (ⅰ) decreases blood pressure and slightly increases hematocrits; (ⅱ) rapidly increases the concentration and urinary excretion of sodium and chloride (≥30-fold), the urine volume (~10-fold), the potassium excretion (two-fold); (ⅲ) contains a potent natriuretic and chloriuretic factor, which strongly inhibits the renal tubular NaCI reabsorption. |
| ANP; Can extracts from other sources induce a natriuretic and diuretic response? (de Bold AJ and Salerno TA, 1983)[3] | Natriuresis and diuresis is induced by atrial extracts from all mammalian species, frog atrial and ventricular extracts, hen ventricle extracts (only diuresis), and not by hen atrial extracts or rat tissue extracts other than the atrial extract; (ⅱ) Natriuretic activity is restricted to heart. |
| ANP; What are the molecules responsible for these activities? (de Bold AJ and Flynn TG, 1983)[4] | Cardionatrin I, which also has effect on vascular smooth muscle tone, has a molecular mass of 5.1 kDa by urea-SDS-PAGE and 49 amino acid residues one of which is cysteine. |
| ANP; What is the common precursor of Cardionatrin I and other atrial peptides? (Flynn TG et al., 1985)[5] | (ⅰ) Cardionatrin IV, consisting of 126 amino acids, has a molecular mass of 19 kDa by urea-SDS-PAGE, and begins immediately after the signal peptide sequence of procardionatrin at residue 25. It does not contain residues 151 and 152, which are arginines; (ⅱ) Cardionatrin III begins at residue 73 and Cardionatrin I begins at residue 123; (ⅲ) Cardionatrins I, III, many cleavage fragments thereof and numerous versions of the carboxyl terminal portion of Cardionatrin I are products derived from a common precursor, Cardionatrin IV; (ⅳ) Cardionatrins I-IV peptides are derived from preprocardionatrin, a common precursor of 152 amino acids (in the rat); (ⅴ) The biologically active sequences of the atrial natriuretic factor is contained in the COOH terminal portion of the molecule. |
| BNP; Identification in porcine brain of a novel natriuretic peptide (Sudoh T et al., 1988)[10] | BNP contains 26 amino acid residues, two Cys residues, seven amino acid substitutions and one addition of (Arg) compared to α-ANP; (ⅱ) BNP possesses diuretic-natriuretic (e.g., increase in urine output, Na+, K+, Cl– excretion) and hypotensive (decrease in mean blood pressure) responses similar to that of ANP; (ⅲ) There may be a dual mechanism involving both ANP and BNP to control physiological functions such as water intake and salt appetite. |
| BNP; What is the intracellular localization of BNP in human cardiac myocytes? (Nakamura S et al., 1991)[11] | BNP is specifically localized in only some of the secretory granules of the human atrium and ventricle that contain ANP, as shown with different patients (with aortic regurgitation, mitral regurgitation or autoptic); (ⅱ) The atrium is the major production site of BNP; (ⅲ) Together, ANP and BNP allows the human heart to regulate blood pressure and body fluid. |
| ANP and BNP; Natriuretic peptides circulate in blood (Clerico et al., 2011 and citations therein)[12] | (ⅰ) Posttranslational processing of proBNP is required for secretion and bioactivity- this process is impaired in patients with heart failure leading to biologically inactive BNP; (ⅱ) proBNP-derived fragments (e.g., the intact and glycosylated forms of precursor proBNP, NH2-terminal-truncated BNP form 3-32) circulate in human plasma in addition to bioactive BNP1-32; (ⅲ) In plasma of patients with heart failure, a significant portion of immunoreactive B-type related peptides is comprised of intact or glycosylated forms of proBNP this suggests that plasma proteases cleave the circulating proBNP to produce biologically active BNP; (ⅳ) In experimental models and in patients with chronic heart failure, a resistance to ANP and BNP is observed; possible mechanisms of resistance to biological effects of ANP and BNP may operate at: a) the pre-receptor level (e.g., existence of inactive natriuretic peptides in plasma, increase in inactivation and degradation of active natriuretic peptides, decreased renal filtration), b) receptor level (e.g., downregulation of NPR-A and NPR-B in target tissues, altered ANP/BNP receptor binding or desensitization, or c) post-receptor level (e.g., altered intracellular signaling). |
| CNP; Are cardiomyocytes able to produce CNP? (Del Ry S et al., 2011)[13] | (ⅰ) Both HUVEC and H9c2 muscle cells express CNP (150 and 200 bp); which can be confirmed in neonatal rat primary cardiomyocytes; (ⅱ) CNP can be immunodetected in both H9c2 cells (by radioimmunologic assay) and cardiomyocytes of pig hearts; (ⅲ) CNP is constitutively expressed in cardiomyocytes. |
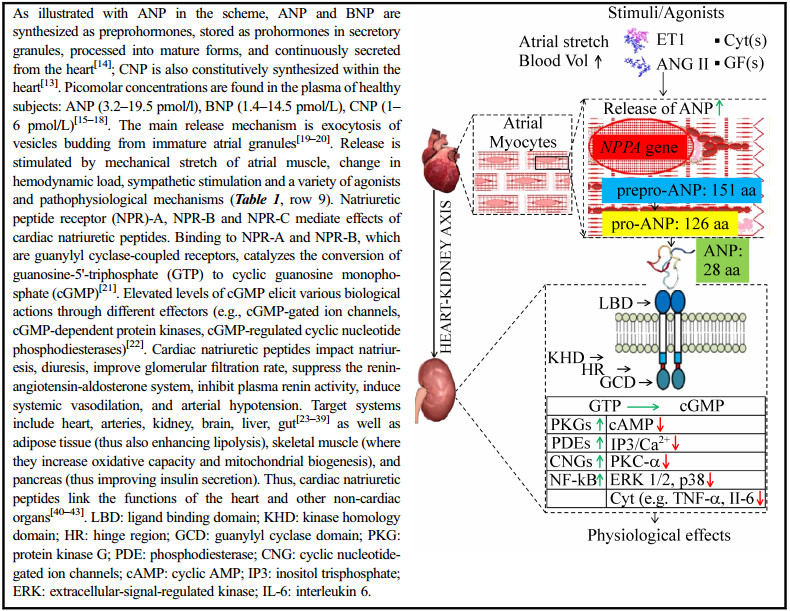
| ANP and BNP; Biological factors and pathophysiological mechanisms that stimulate the production /release of natriuretic peptides (Clerico et al., 2011 and citations therein)[12] | (ⅰ) The production/release of cNPs is stimulated by: a) Ang II, ET1, α-adrenergic agonists, cytokines such as IL-1, IL-6 and TNF-α, and lipopolysaccharide (all of which signal throughout NF-kB activated by MAPK), b) arginine vasopressin (through Ca2+ influx and PKC), c) GFs (signaling through MAPK cascade), d) prostaglandins (through PLC, IP3, PKC, and MLCK), e) chromogranin B (through NF-kB and IP3/ Ca2+ influx), f) thyroid hormones (through thyroid hormone regulatory element), g) corticosteroids (through glucocorticoid responsive element), and h) estrogens; (ⅱ) The production and release of BNP from ventricular cardiomyocytes is stimulated by inflammation, ventricular hypertrophy, and fibrosis; (ⅲ) Even in isolated and cultured ventricular cells, myocardial ischemia can induce the synthesis/secretion of BNP and its related peptides; (ⅳ) Both ANP and BNP transcription may be activated by the hypoxia-inducible factor-1a (which is induced under low oxygen conditions). |
| Cardiac specific miR-208a and MED13; How does cardiac MED13 influence whole body metabolism? (Grueter CE et al., 2012)[6] | (ⅰ) Pharmacologic inhibition of the cardiac-specific miR-208a confers resistance to diet-induced obesity (e.g., smaller visceral WAT and subscapular BAT, normal glucose response, lower fasting insulin levels) with beneficial metabolic effects (e.g., reduced serum triglyceride and cholesterol levels); (ⅱ) miR-208a is a negative transcriptional regulator of MED13 in the heart. Among the functions of MED13 are: (a) to inhibit expression of metabolic genes regulated by NRs (e.g., Gpd2, Thrsp, Cidea, Elovl6, Eno1, PPARγ Tkt), (b) to control whole-body metabolic homeostasis (e.g., αMHC-Med13 TG mice, with increased cardiac expression of MED13, show enhanced metabolic rate, diminished serum triglyceride and cholesterol levels, resistance to diet-induced obesity including less fat mass versus WT littermates, reduced visceral WAT and subscapular BAT mass as well as less adipocyte size and less lipid accumulation, improved glucose (tolerance) response, lowered plasma lipid levels, and improved whole-body insulin sensitivity, and (c) regulate energy expenditure (e.g., increased oxygen consumption, carbon dioxide production) in mice; (ⅲ) MED13 deficiency in the heart increases susceptibility to metabolic syndrome and diet-induced obesity in mice, as shown with Med13 cardiac knockout mice versus Med13fl/fl littermates on HF diet. (ⅳ) Circulating factors may relay MED13 activity from the heart to other organs but these factors remain elusive. |
| Abbreviations: miR-208a: microRNA-208a; MED13: mediator complex subunit 13; ANP: atrial natriuretic peptide; SDS-PAGE: sodium dodecyl sulfate-polyacrylamide gel electrophoresis; BNP: brain natriuretic peptide; proBNP: ventricular circulating inactive precursor of BNP; CNP: C-type natriuretic peptide; HUVEC: human umbilical vein endothelial cell; NRs: nuclear receptors; TG: transgenic; Gpd2: glycerol-3-phosphate dehydrogenase 2; Thrsp: thyroid hormone responsive; Cidea: cell death-inducing DFFA-like effector A; Elovl6: ELOVL fatty acid elongase 6; Eno1: Enolase 1; PPARγ: peroxisome proliferator activated receptor gamma; Tkt: Transketolase; WT: wild-type; WAT: white adipose tissue; BAT: brown adipose tissue; HF: high fat; Ang II: angiotensin II; ET1: endothelin-1; NF-kB: nuclear factor kappa-light-chain-enhancer of activated B cells; MAPK: p38 mitogen-activated protein kinase; PKC: protein kinase C; GFs: growth factors; MLCK: myosin light chain kinase; PLC: phospholipase C. | |
 DownLoad:
DownLoad: