| [1] |
Chakraborty S, Jasnin M, Baumeister W. Three-dimensional organization of the cytoskeleton: a cryo-electron tomography perspective[J]. Protein Sci, 2020, 29(6): 1302–1320. doi: 10.1002/pro.3858
|
| [2] |
Lorenz C, Köster S. Multiscale architecture: mechanics of composite cytoskeletal networks[J]. Biophys Rev (Melville), 2022, 3(3): 031304. doi: 10.1063/5.0099405
|
| [3] |
Doganyigit Z, Eroglu E, Okan A. Intermediate filament proteins are reliable immunohistological biomarkers to help diagnose multiple tissue-specific diseases[J]. Anat Histol Embryol, 2023, 52(5): 655–672. doi: 10.1111/ahe.12937
|
| [4] |
Barritt J, King AT, Pickard JN. The effects of cystine diet on keratin composition in rabbit wool[J]. Biochem J, 1930, 24(4): 1061–1065. doi: 10.1042/bj0241061
|
| [5] |
Palay SL, Palade GE. The fine structure of neurons[J]. J Biophys Biochem Cytol, 1955, 1(1): 69–88. doi: 10.1083/jcb.1.1.69
|
| [6] |
Ishikawa H, Bischoff R, Holtzer H. Formation of arrowhead complexes with heavy meromyosin in a variety of cell types[J]. J Cell Biol, 1969, 43(2): 312–328. doi: 10.1083/jcb.43.2.312
|
| [7] |
Franke WW, Schmid E, Osborn M, et al. Different intermediate-sized filaments distinguished by immunofluorescence microscopy[J]. Proc Natl Acad Sci U S A, 1978, 75(10): 5034–5038. doi: 10.1073/pnas.75.10.5034
|
| [8] |
Schmid E, Tapscott S, Bennett GS, et al. Differential location of different types of intermediate-sized filaments in various tissues of the chicken embryo[J]. Differentiation, 1979, 15(1): 27–40. https://pubmed.ncbi.nlm.nih.gov/93557/
|
| [9] |
Goldmann WH. Intermediate filaments and cellular mechanics[J]. Cell Biol Int, 2018, 42(2): 132–138. doi: 10.1002/cbin.10879
|
| [10] |
Gao J, Nakamura F. Actin-associated proteins and small molecules targeting the actin cytoskeleton[J]. Int J Mol Sci, 2022, 23(4): 2118. doi: 10.3390/ijms23042118
|
| [11] |
Peng N, Nakamura F. Microtubule-associated proteins and enzymes modifying tubulin[J]. Cytoskeleton (Hoboken), 2023, 80(3-4): 60–76. doi: 10.1002/cm.21748
|
| [12] |
Bott CJ, Winckler B. Intermediate filaments in developing neurons: beyond structure[J]. Cytoskeleton (Hoboken), 2020, 77(3-4): 110–128. doi: 10.1002/cm.21597
|
| [13] |
Dutour-Provenzano G, Etienne-Manneville S. Intermediate filaments[J]. Curr Biol, 2021, 31(10): R522–R529. doi: 10.1016/j.cub.2021.04.011
|
| [14] |
Eldirany SA, Lomakin IB, Ho M, et al. Recent insight into intermediate filament structure[J]. Curr Opin Cell Biol, 2021, 68: 132–143. doi: 10.1016/j.ceb.2020.10.001
|
| [15] |
Sjöqvist M, Antfolk D, Suarez-Rodriguez F, et al. From structural resilience to cell specification-intermediate filaments as regulators of cell fate[J]. FASEB J, 2021, 35(1): e21182. https://pubmed.ncbi.nlm.nih.gov/33205514/
|
| [16] |
Romano R, Del Fiore VS, Bucci C. Role of the intermediate filament protein peripherin in health and disease[J]. Int J Mol Sci, 2022, 23(23): 15416. doi: 10.3390/ijms232315416
|
| [17] |
Infante E, Etienne-Manneville S. Intermediate filaments: integration of cell mechanical properties during migration[J]. Front Cell Dev Biol, 2022, 10: 951816. doi: 10.3389/fcell.2022.951816
|
| [18] |
Coulombe PA, Pineda CM, Jacob JT, et al. Nuclear roles for non-lamin intermediate filament proteins[J]. Curr Opin Cell Biol, 2024, 86: 102303. doi: 10.1016/j.ceb.2023.102303
|
| [19] |
Pogoda K, Janmey PA. Transmit and protect: the mechanical functions of intermediate filaments[J]. Curr Opin Cell Biol, 2023, 85: 102281. doi: 10.1016/j.ceb.2023.102281
|
| [20] |
Schwarz N, Leube RE. Plasticity of cytoplasmic intermediate filament architecture determines cellular functions[J]. Curr Opin Cell Biol, 2023, 85: 102270. doi: 10.1016/j.ceb.2023.102270
|
| [21] |
Utsunomiya H, Fujita M, Naito F, et al. Cell cycle-dependent dynamics of a plant intermediate filament motif protein with intracellular localization related to microtubules[J]. Protoplasma, 2020, 257(5): 1387–1400. doi: 10.1007/s00709-020-01512-1
|
| [22] |
Shymanovich T, Vandenbrink JP, Herranz R, et al. Spaceflight studies identify a gene encoding an intermediate filament involved in tropism pathways[J]. Plant Physiol Biochem, 2022, 171: 191–200. doi: 10.1016/j.plaphy.2021.12.039
|
| [23] |
Jacob JT, Coulombe PA, Kwan R, et al. Types Ⅰ and Ⅱ keratin intermediate filaments[J]. Cold Spring Harb Perspect Biol, 2018, 10(4): a018275. doi: 10.1101/cshperspect.a018275
|
| [24] |
Mohamad J, Sarig O, Beattie P, et al. A unique skin phenotype resulting from a large heterozygous deletion spanning six keratin genes[J]. Br J Dermatol, 2022, 187(5): 773–777. doi: 10.1111/bjd.21766
|
| [25] |
Li P, Rietscher K, Jopp H, et al. Posttranslational modifications of keratins and their associated proteins as therapeutic targets in keratin diseases[J]. Curr Opin Cell Biol, 2023, 85: 102264. doi: 10.1016/j.ceb.2023.102264
|
| [26] |
|
| [27] |
Walko G, Castañón MJ, Wiche G. Molecular architecture and function of the hemidesmosome[J]. Cell Tissue Res, 2015, 360(3): 529–544. doi: 10.1007/s00441-015-2216-6
|
| [28] |
Evtushenko NA, Beilin AK, Kosykh AV, et al. Keratins as an inflammation trigger point in epidermolysis bullosa simplex[J]. Int J Mol Sci, 2021, 22(22): 12446. doi: 10.3390/ijms222212446
|
| [29] |
Chen F, Yao L, Zhang X, et al. Damaged keratin filament network caused by KRT5 mutations in localized recessive epidermolysis bullosa simplex[J]. Front Genet, 2021, 12: 736610. doi: 10.3389/fgene.2021.736610
|
| [30] |
Has C, Fischer J. Inherited epidermolysis bullosa: new diagnostics and new clinical phenotypes[J]. Exp Dermatol, 2019, 28(10): 1146–1152. doi: 10.1111/exd.13668
|
| [31] |
DeStefano GM, Christiano AM. The genetics of human skin disease[J]. Cold Spring Harb Perspect Med, 2014, 4(10): a015172. doi: 10.1101/cshperspect.a015172
|
| [32] |
Leube RE, Schwarz N. Current mysteries of pachyonychia congenita[J]. Br J Dermatol, 2020, 182(3): 525–526. doi: 10.1111/bjd.18688
|
| [33] |
Goldfarb LG, Olivé M, Vicart P, et al. Intermediate filament diseases: desminopathy[M]//Laing NG. The Sarcomere and Skeletal Muscle Disease. New York: Springer, 2008: 131–164.
|
| [34] |
Kubánek M, Schimerová T, Piherová L, et al. Desminopathy: novel desmin variants, a new cardiac phenotype, and further evidence for secondary mitochondrial dysfunction[J]. J Clin Med, 2020, 9(4): 937. doi: 10.3390/jcm9040937
|
| [35] |
Su W, van Wijk SW, Brundel BJJM. Desmin variants: trigger for cardiac arrhythmias?[J]. Front Cell Dev Biol, 2022, 10: 986718. doi: 10.3389/fcell.2022.986718
|
| [36] |
Marunouchi T, Inomata S, Sanbe A, et al. Protective effect of geranylgeranylacetone via enhanced induction of HSPB1 and HSPB8 in mitochondria of the failing heart following myocardial infarction in rats[J]. Eur J Pharmacol, 2014, 730: 140–147. doi: 10.1016/j.ejphar.2014.02.037
|
| [37] |
Sanbe A, Daicho T, Mizutani R, et al. Protective effect of geranylgeranylacetone via enhancement of HSPB8 induction in desmin-related cardiomyopathy[J]. PLoS One, 2009, 4(4): e5351. doi: 10.1371/journal.pone.0005351
|
| [38] |
Viedma-Poyatos Á, Pajares MA, Pérez-Sala D. Type Ⅲ intermediate filaments as targets and effectors of electrophiles and oxidants[J]. Redox Biol, 2020, 36: 101582. doi: 10.1016/j.redox.2020.101582
|
| [39] |
Pérez-Sala D, Quinlan RA. The redox-responsive roles of intermediate filaments in cellular stress detection, integration and mitigation[J]. Curr Opin Cell Biol, 2024, 86: 102283. doi: 10.1016/j.ceb.2023.102283
|
| [40] |
Botha CJ, Mathe YZ, Ferreira GCH, et al. Cytotoxicity of the sesquiterpene lactones, ivalin and parthenolide in murine muscle cell lines and their effect on desmin, a cytoskeletal intermediate filament[J]. Toxins (Basel), 2020, 12(7): 459. doi: 10.3390/toxins12070459
|
| [41] |
|
| [42] |
Jaramillo-Rangel G, Chávez-Briones MDL, Ancer-Arellano A, et al. Nestin-expressing cells in the lung: the bad and the good parts[J]. Cells, 2021, 10(12): 3413. doi: 10.3390/cells10123413
|
| [43] |
Maggi L, Mavroidis M, Psarras S, et al. Skeletal and cardiac muscle disorders caused by mutations in genes encoding intermediate filament proteins[J]. Int J Mol Sci, 2021, 22(8): 4256. doi: 10.3390/ijms22084256
|
| [44] |
Ducray F, Criniere E, Idbaih A, et al. α-Internexin expression identifies 1p19q codeleted gliomas[J]. Neurology, 2009, 72(2): 156–161. doi: 10.1212/01.wnl.0000339055.64476.cb
|
| [45] |
Yuan A, Rao MV, Veeranna, et al. Neurofilaments and neurofilament proteins in health and disease[J]. Cold Spring Harb Perspect Biol, 2017, 9(4): a018309. doi: 10.1101/cshperspect.a018309
|
| [46] |
Kotaich F, Caillol D, Bomont P. Neurofilaments in health and Charcot-Marie-Tooth disease[J]. Front Cell Dev Biol, 2023, 11: 1275155. doi: 10.3389/fcell.2023.1275155
|
| [47] |
Stone EJ, Kolb SJ, Brown A. A review and analysis of the clinical literature on Charcot-Marie-Tooth disease caused by mutations in neurofilament protein L[J]. Cytoskeleton (Hoboken), 2021, 78(3): 97–110. doi: 10.1002/cm.21676
|
| [48] |
|
| [49] |
Cortese A, Wilcox JE, Polke JM, et al. Targeted next-generation sequencing panels in the diagnosis of Charcot-Marie-Tooth disease[J]. Neurology, 2020, 94(1): e51–e61. https://pubmed.ncbi.nlm.nih.gov/31827005/
|
| [50] |
Stone EJ, Uchida A, Brown A. Charcot-Marie-Tooth disease Type 2E/1F mutant neurofilament proteins assemble into neurofilaments[J]. Cytoskeleton (Hoboken), 2019, 76(7-8): 423–439. doi: 10.1002/cm.21566
|
| [51] |
Nowogrodzka K, Jankowska-Konsur A. Emerging biomarker in carcinogenesis. Focus on nestin[J]. Postepy Dermatol Alergol, 2022, 39(6): 1001–1007. doi: 10.5114/ada.2022.122599
|
| [52] |
Russell MA. Synemin redefined: multiple binding partners results in multifunctionality[J]. Front Cell Dev Biol, 2020, 8: 159. doi: 10.3389/fcell.2020.00159
|
| [53] |
Paulin D, Hovhannisyan Y, Kasakyan S, et al. Synemin-related skeletal and cardiac myopathies: an overview of pathogenic variants[J]. Am J Physiol Cell Physiol, 2020, 318(4): C709–C718. doi: 10.1152/ajpcell.00485.2019
|
| [54] |
Song S, Landsbury A, Dahm R, et al. Functions of the intermediate filament cytoskeleton in the eye lens[J]. J Clin Invest, 2009, 119(7): 1837–1848. doi: 10.1172/JCI38277
|
| [55] |
Wang H, Zhang T, Wu D, et al. A novel beaded filament structural protein 1 ( BFSP1) gene mutation associated with autosomal dominant congenital cataract in a Chinese family[J]. Mol Vis, 2013, 19: 2590–2595. https://pubmed.ncbi.nlm.nih.gov/24379646/
|
| [56] |
Liu Q, Wang K, Zhu S. A novel p.G112E mutation in BFSP2 associated with autosomal dominant pulverulent cataract with sutural opacities[J]. Curr Eye Res, 2014, 39(10): 1013–1019. doi: 10.3109/02713683.2014.891749
|
| [57] |
Wang H, Ouyang G, Zhu Y. D348N mutation of BFSP1 gene in congenital cataract: it does matter[J]. Cell Biochem Biophys, 2023, 81(4): 757–763. doi: 10.1007/s12013-023-01169-6
|
| [58] |
Cvekl A, Camerino MJ. Generation of lens progenitor cells and lentoid bodies from pluripotent stem cells: novel tools for human lens development and ocular disease etiology[J]. Cells, 2022, 11(21): 3516. doi: 10.3390/cells11213516
|
| [59] |
Marcelot A, Worman HJ, Zinn-Justin S. Protein structural and mechanistic basis of progeroid laminopathies[J]. FEBS J, 2021, 288(9): 2757–2772. doi: 10.1111/febs.15526
|
| [60] |
Shah PP, Santini GT, Shen KM, et al. InterLINCing chromatin organization and mechanobiology in laminopathies[J]. Curr Cardiol Rep, 2023, 25(5): 307–314. doi: 10.1007/s11886-023-01853-2
|
| [61] |
Wong X, Stewart CL. The laminopathies and the insights they provide into the structural and functional organization of the nucleus[J]. Annu Rev Genomics Hum Genet, 2020, 21: 263–288. doi: 10.1146/annurev-genom-121219-083616
|
| [62] |
Malashicheva A, Perepelina K. Diversity of nuclear lamin A/C action as a key to tissue-specific regulation of cellular identity in health and disease[J]. Front Cell Dev Biol, 2021, 9: 761469. doi: 10.3389/fcell.2021.761469
|
| [63] |
Yamada S, Ko T, Ito M, et al. TEAD1 trapping by the Q353R-Lamin A/C causes dilated cardiomyopathy[J]. Sci Adv, 2023, 9(15): eade7047. doi: 10.1126/sciadv.ade7047
|
| [64] |
|
| [65] |
Evangelisti C, Rusciano I, Mongiorgi S, et al. The wide and growing range of lamin B-related diseases: from laminopathies to cancer[J]. Cell Mol Life Sci, 2022, 79(2): 126. doi: 10.1007/s00018-021-04084-2
|
| [66] |
Samen U, Eikmanns BJ, Reinscheid DJ, et al. The surface protein Srr-1 of Streptococcus agalactiae binds human keratin 4 and promotes adherence to epithelial HEp-2 cells[J]. Infect Immun, 2007, 75(11): 5405–5414. doi: 10.1128/IAI.00717-07
|
| [67] |
Das S, Ravi V, Desai A. Japanese encephalitis virus interacts with vimentin to facilitate its entry into porcine kidney cell line[J]. Virus Res, 2011, 160(1-2): 404–408. doi: 10.1016/j.virusres.2011.06.001
|
| [68] |
Deng L, Spencer BL, Holmes JA, et al. The Group B Streptococcal surface antigen Ⅰ/Ⅱ protein, BspC, interacts with host vimentin to promote adherence to brain endothelium and inflammation during the pathogenesis of meningitis[J]. PLoS Pathog, 2019, 15(6): e1007848. doi: 10.1371/journal.ppat.1007848
|
| [69] |
Ma X, Ling Y, Li P, et al. Cellular vimentin interacts with foot-and-mouth disease virus nonstructural protein 3A and negatively modulates viral replication[J]. J Virol, 2020, 94(16): e00273–20. https://pubmed.ncbi.nlm.nih.gov/32493819/
|
| [70] |
Wang A, Liu X, Heckmann A, et al. A Trichinella spiralis new born larvae-specific protein, Ts-NBL1, interacts with host's cell vimentin[J]. Parasitol Res, 2022, 121(5): 1369–1378. doi: 10.1007/s00436-022-07479-7
|
| [71] |
Deptuła P, Fiedoruk K, Wasilewska M, et al. Physicochemical nature of SARS-CoV-2 spike protein binding to human vimentin[J]. ACS Appl Mater Interfaces, 2023, 15(28): 34172–34180. doi: 10.1021/acsami.3c03347
|
| [72] |
Zhang Y, Zhao S, Li Y, et al. Host cytoskeletal vimentin serves as a structural organizer and an RNA-binding protein regulator to facilitate Zika viral replication[J]. Proc Natl Acad Sci U S A, 2022, 119(8): e2113909119. doi: 10.1073/pnas.2113909119
|
| [73] |
Risinger AL, Du L. Targeting and extending the eukaryotic druggable genome with natural products: cytoskeletal targets of natural products[J]. Nat Prod Rep, 2020, 37(5): 634–652. doi: 10.1039/C9NP00053D
|
| [74] |
Kerns ML, DePianto D, Dinkova-Kostova AT, et al. Reprogramming of keratin biosynthesis by sulforaphane restores skin integrity in epidermolysis bullosa simplex[J]. Proc Natl Acad Sci U S A, 2007, 104(36): 14460–14465. doi: 10.1073/pnas.0706486104
|
| [75] |
Virtanen M, Gedde-Dahl T Jr, Mörk NJ, et al. Phenotypic/genotypic correlations in patients with epidermolytic hyperkeratosis and the effects of retinoid therapy on keratin expression[J]. Acta Derm Venereol, 2001, 81(3): 163–170. doi: 10.1080/000155501750376221
|
| [76] |
Zieman AG, Poll BG, Ma J, et al. Altered keratinocyte differentiation is an early driver of keratin mutation-based palmoplantar keratoderma[J]. Hum Mol Genet, 2019, 28(13): 2255–2270. doi: 10.1093/hmg/ddz050
|
| [77] |
Kim KH, Jung JH, Chung WS, et al. Ferulic acid induces keratin 6α via inhibition of nuclear β-catenin accumulation and activation of Nrf2 in wound-induced inflammation[J]. Biomedicines, 2021, 9(5): 459. doi: 10.3390/biomedicines9050459
|
| [78] |
Kwan R, Looi K, Omary MB. Absence of keratins 8 and 18 in rodent epithelial cell lines associates with keratin gene mutation and DNA methylation: cell line selective effects on cell invasion[J]. Exp Cell Res, 2015, 335(1): 12–22. doi: 10.1016/j.yexcr.2015.04.003
|
| [79] |
Rietscher K, Jahnke HG, Rübsam M, et al. Kinase inhibition by PKC412 prevents epithelial sheet damage in autosomal dominant epidermolysis bullosa simplex through keratin and cell contact stabilization[J]. J Invest Dermatol, 2022, 142(12): 3282–3293. doi: 10.1016/j.jid.2022.05.1088
|
| [80] |
Ziaei E, de Paiva IM, Yao SJ, et al. Peptide-drug conjugate targeting keratin 1 inhibits triple-negative breast cancer in mice[J]. Mol Pharm, 2023, 20(7): 3570–3577. doi: 10.1021/acs.molpharmaceut.3c00189
|
| [81] |
Lee GH, Lekwuttikarn R, Tafoya E, et al. Transcriptomic repositioning analysis identifies mTOR inhibitor as potential therapy for epidermolysis bullosa simplex[J]. J Invest Dermatol, 2022, 142(2): 382–389. doi: 10.1016/j.jid.2021.07.170
|
| [82] |
Cabet E, Batonnet-Pichon S, Delort F, et al. Antioxidant treatment and induction of autophagy cooperate to reduce desmin aggregation in a cellular model of desminopathy[J]. PLoS One, 2015, 10(9): e0137009. doi: 10.1371/journal.pone.0137009
|
| [83] |
Bachetti T, Zanni ED, Adamo A, et al. Beneficial effect of phenytoin and carbamazepine on GFAP gene expression and mutant GFAP folding in a cellular model of Alexander's disease[J]. Front Pharmacol, 2021, 12: 723218. doi: 10.3389/fphar.2021.723218
|
| [84] |
Satelli A, Li S. Vimentin in cancer and its potential as a molecular target for cancer therapy[J]. Cell Mol Life Sci, 2011, 68(18): 3033–3046. doi: 10.1007/s00018-011-0735-1
|
| [85] |
Bollong MJ, Pietilä M, Pearson AD, et al. A vimentin binding small molecule leads to mitotic disruption in mesenchymal cancers[J]. Proc Natl Acad Sci U S A, 2017, 114(46): E9903–E9912. doi: 10.1073/pnas.1716009114
|
| [86] |
Ramos I, Stamatakis K, Oeste CL, et al. Vimentin as a multifaceted player and potential therapeutic target in viral infections[J]. Int J Mol Sci, 2020, 21(13): 4675. doi: 10.3390/ijms21134675
|
| [87] |
Bargagna-Mohan P, Hamza A, Kim YE, et al. The tumor inhibitor and antiangiogenic agent Withaferin A targets the intermediate filament protein vimentin[J]. Chem Biol, 2007, 14(6): 623–634. doi: 10.1016/j.chembiol.2007.04.010
|
| [88] |
|
| [89] |
Bargagna-Mohan P, Paranthan RR, Hamza A, et al. Withaferin A targets intermediate filaments glial fibrillary acidic protein and vimentin in a model of retinal gliosis[J]. J Biol Chem, 2010, 285(10): 7657–7669. doi: 10.1074/jbc.M109.093765
|
| [90] |
Bargagna-Mohan P, Paranthan RR, Hamza A, et al. Corneal antifibrotic switch identified in genetic and pharmacological deficiency of vimentin[J]. J Biol Chem, 2012, 287(2): 989–1006. doi: 10.1074/jbc.M111.297150
|
| [91] |
Thaiparambil JT, Bender L, Ganesh T, et al. Withaferin A inhibits breast cancer invasion and metastasis at sub-cytotoxic doses by inducing vimentin disassembly and serine 56 phosphorylation[J]. Int J Cancer, 2011, 129(11): 2744–2755. doi: 10.1002/ijc.25938
|
| [92] |
de Pablo Y, Chen M, Möllerström E, et al. Drugs targeting intermediate filaments can improve neurosupportive properties of astrocytes[J]. Brain Res Bull, 2018, 136: 130–138. doi: 10.1016/j.brainresbull.2017.01.021
|
| [93] |
Kaschula CH, Tuveri R, Ngarande E, et al. The garlic compound ajoene covalently binds vimentin, disrupts the vimentin network and exerts anti-metastatic activity in cancer cells[J]. BMC Cancer, 2019, 19(1): 248. doi: 10.1186/s12885-019-5388-8
|
| [94] |
Trogden KP, Battaglia RA, Kabiraj P, et al. An image-based small-molecule screen identifies vimentin as a pharmacologically relevant target of simvastatin in cancer cells[J]. FASEB J, 2018, 32(5): 2841–2854. doi: 10.1096/fj.201700663R
|
| [95] |
Kim HR, Warrington SJ, López-Guajardo A, et al. ALD-R491 regulates vimentin filament stability and solubility, cell contractile force, cell migration speed and directionality[J]. Front Cell Dev Biol, 2022, 10: 926283. doi: 10.3389/fcell.2022.926283
|
| [96] |
Rezaeianpour M, Mazidi SM, Nami R, et al. Vimentin-targeted radiopeptide 99mTc-HYNIC-(tricine/EDDA)-VNTANST: a promising drug for pulmonary fibrosis imaging[J]. Nucl Med Commun, 2023, 44(9): 777–787. doi: 10.1097/MNM.0000000000001724
|
| [97] |
He S, Lin J, Lin L, et al. Shikonin-mediated inhibition of nestin affects hypoxia-induced proliferation of pulmonary artery smooth muscle cells[J]. Mol Med Rep, 2018, 18(3): 3476–3482. https://pubmed.ncbi.nlm.nih.gov/30066896/
|
| [98] |
Feng X, Han H, Guo Y, et al. LncRNA ENST869 targeting Nestin transcriptional region to affect the pharmacological effects of chidamide in breast cancer cells[J]. Front Oncol, 2022, 12: 874343. doi: 10.3389/fonc.2022.874343
|
| [99] |
Lee SJ, Jung YS, Yoon MH, et al. Interruption of progerin-lamin A/C binding ameliorates Hutchinson-Gilford progeria syndrome phenotype[J]. J Clin Invest, 2016, 126(10): 3879–3893. doi: 10.1172/JCI84164
|
| [100] |
Kang SM, Yoon MH, Ahn J, et al. Progerinin, an optimized progerin-lamin A binding inhibitor, ameliorates premature senescence phenotypes of Hutchinson-Gilford progeria syndrome[J]. Commun Biol, 2021, 4(1): 5. doi: 10.1038/s42003-020-01540-w
|
| [101] |
Kang SM, Seo S, Song EJ, et al. Progerinin, an inhibitor of progerin, alleviates cardiac abnormalities in a model mouse of Hutchinson-Gilford progeria syndrome[J]. Cells, 2023, 12(9): 1232. doi: 10.3390/cells12091232
|
| [102] |
Glynn MW, Glover TW. Incomplete processing of mutant lamin A in Hutchinson-Gilford progeria leads to nuclear abnormalities, which are reversed by farnesyltransferase inhibition[J]. Hum Mol Genet, 2005, 14(20): 2959–2969. doi: 10.1093/hmg/ddi326
|
| [103] |
Finley J. Alteration of splice site selection in the LMNA gene and inhibition of progerin production via AMPK activation[J]. Med Hypotheses, 2014, 83(5): 580–587. doi: 10.1016/j.mehy.2014.08.016
|
| [104] |
Finley J. Cellular stress and AMPK activation as a common mechanism of action linking the effects of metformin and diverse compounds that alleviate accelerated aging defects in Hutchinson-Gilford progeria syndrome[J]. Med Hypotheses, 2018, 118: 151–162. doi: 10.1016/j.mehy.2018.06.029
|
| [105] |
Kim BH, Woo TG, Kang SM, et al. Splicing variants, protein-protein interactions, and drug targeting in Hutchinson-Gilford progeria syndrome and small cell lung cancer[J]. Genes (Basel), 2022, 13(2): 165. doi: 10.3390/genes13020165
|
| [106] |
|
| [107] |
Tsai CF, Wu JY, Hsu YW. Protective effects of rosmarinic acid against selenite-induced cataract and oxidative damage in rats[J]. Int J Med Sci, 2019, 16(5): 729–740. doi: 10.7150/ijms.32222
|
| [108] |
Janmey PA, Euteneuer U, Traub P, et al. Viscoelastic properties of vimentin compared with other filamentous biopolymer networks[J]. J Cell Biol, 1991, 113(1): 155–160. doi: 10.1083/jcb.113.1.155
|
| [109] |
Huber F, Boire A, López MP, et al. Cytoskeletal crosstalk: when three different personalities team up[J]. Curr Opin Cell Biol, 2015, 32(24): 39–47. doi: 10.1016/j.ceb.2014.10.005
|
| [110] |
Rölleke U, Kumari P, Meyer R, et al. The unique biomechanics of intermediate filaments – from single filaments to cells and tissues[J]. Curr Opin Cell Biol, 2023, 85: 102263. doi: 10.1016/j.ceb.2023.102263
|
| [111] |
Wen Q, Janmey PA. Polymer physics of the cytoskeleton[J]. Curr Opin Solid State Mater Sci, 2011, 15(5): 177–182. doi: 10.1016/j.cossms.2011.05.002
|
| [112] |
Sapra KT, Medalia O. Bend, push, stretch: remarkable structure and mechanics of single intermediate filaments and meshworks[J]. Cells, 2021, 10(8): 1960. doi: 10.3390/cells10081960
|
| [113] |
van Bodegraven EJ, Etienne-Manneville S. Intermediate filaments from tissue integrity to single molecule mechanics[J]. Cells, 2021, 10(8): 1905. doi: 10.3390/cells10081905
|
| [114] |
Hu J, Li Y, Hao Y, et al. High stretchability, strength, and toughness of living cells enabled by hyperelastic vimentin intermediate filaments[J]. Proc Natl Acad Sci U S A, 2019, 116(35): 17175–17180. doi: 10.1073/pnas.1903890116
|
| [115] |
Ackbarow T, Sen D, Thaulow C, et al. Alpha-helical protein networks are self-protective and flaw-tolerant[J]. PLoS One, 2009, 4(6): e6015. doi: 10.1371/journal.pone.0006015
|
| [116] |
Block J, Witt H, Candelli A, et al. Nonlinear loading-rate-dependent force response of individual vimentin intermediate filaments to applied strain[J]. Phys Rev Lett, 2017, 118(4): 048101. doi: 10.1103/PhysRevLett.118.048101
|
| [117] |
Johnson CP, Tang HY, Carag C, et al. Forced unfolding of proteins within cells[J]. Science, 2007, 317(5838): 663–666. doi: 10.1126/science.1139857
|
| [118] |
Fleissner F, Kumar S, Klein N, et al. Tension causes unfolding of intracellular vimentin intermediate filaments[J]. Adv Biosyst, 2020, 4(11): e2000111. doi: 10.1002/adbi.202000111
|
| [119] |
Seltmann K, Fritsch AW, Käs JA, et al. Keratins significantly contribute to cell stiffness and impact invasive behavior[J]. Proc Natl Acad Sci U S A, 2013, 110(46): 18507–18512. doi: 10.1073/pnas.1310493110
|
| [120] |
Ramms L, Fabris G, Windoffer R, et al. Keratins as the main component for the mechanical integrity of keratinocytes[J]. Proc Natl Acad Sci U S A, 2013, 110(46): 18513–18518. doi: 10.1073/pnas.1313491110
|
| [121] |
Charrier EE, Montel L, Asnacios A, et al. The desmin network is a determinant of the cytoplasmic stiffness of myoblasts[J]. Biol Cell, 2018, 110(4): 77–90. doi: 10.1111/boc.201700040
|
| [122] |
Patteson AE, Vahabikashi A, Pogoda K, et al. Vimentin protects cells against nuclear rupture and DNA damage during migration[J]. J Cell Biol, 2019, 218(12): 4079–4092. doi: 10.1083/jcb.201902046
|
| [123] |
Laly AC, Sliogeryte K, Pundel OJ, et al. The keratin network of intermediate filaments regulates keratinocyte rigidity sensing and nuclear mechanotransduction[J]. Sci Adv, 2021, 7(5): eabd6187. doi: 10.1126/sciadv.abd6187
|
| [124] |
Swoger M, Gupta S, Charrier EE, et al. Vimentin intermediate filaments mediate cell morphology on viscoelastic substrates[J]. ACS Appl Bio Mater, 2022, 5(2): 552–561. doi: 10.1021/acsabm.1c01046
|
| [125] |
Alisafaei F, Mandal K, Saldanha R, et al. Vimentin is a key regulator of cell mechanosensing through opposite actions on actomyosin and microtubule networks[J]. Commun Biol, 2024, 7(1): 658. doi: 10.1038/s42003-024-06366-4
|
| [126] |
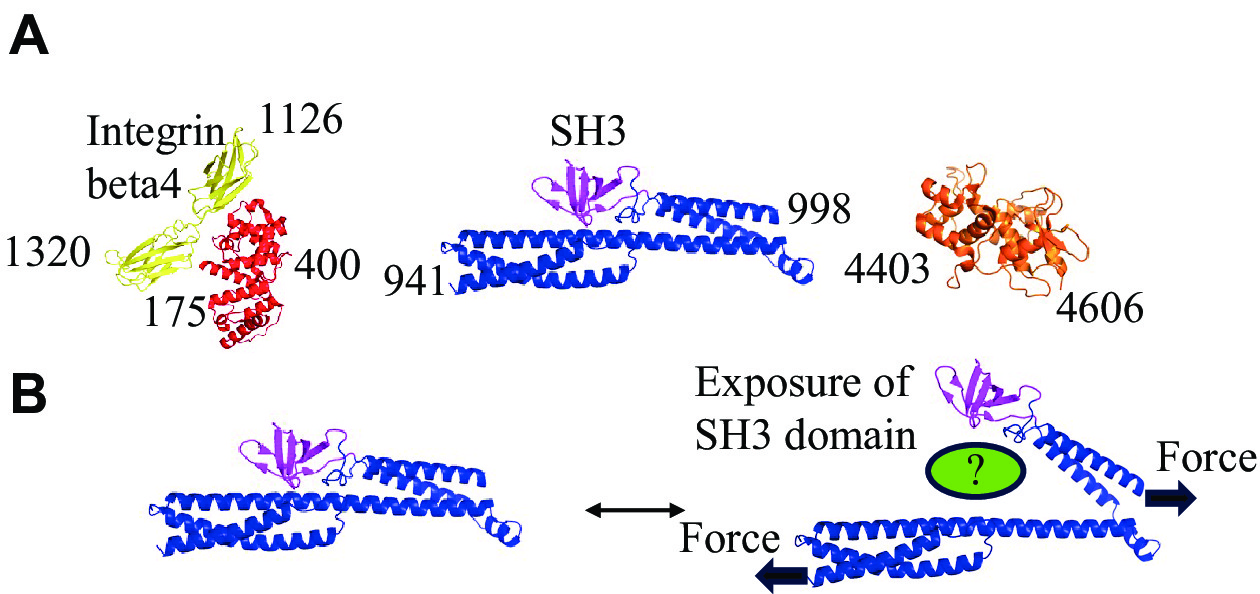
Daday C, Kolšek K, Gräter F. The mechano-sensing role of the unique SH3 insertion in plakin domains revealed by molecular dynamics simulations[J]. Sci Rep, 2017, 7(1): 11669. doi: 10.1038/s41598-017-11017-2
|
| [127] |
Suman SK, Daday C, Ferraro T, et al. The plakin domain of C. elegans VAB-10/plectin acts as a hub in a mechanotransduction pathway to promote morphogenesis[J]. Development, 2019, 146(24): dev183780. doi: 10.1242/dev.183780
|
| [128] |
Na S, Chowdhury F, Tay B, et al. Plectin contributes to mechanical properties of living cells[J]. Am J Physiol Cell Physiol, 2009, 296(4): C868–C877. doi: 10.1152/ajpcell.00604.2008
|
| [129] |
Almeida FV, Walko G, McMillan JR, et al. The cytolinker plectin regulates nuclear mechanotransduction in keratinocytes[J]. J Cell Sci, 2015, 128(24): 4475–4486. doi: 10.1242/jcs.173435
|
| [130] |
Wintner O, Hirsch-Attas N, Schlossberg M, et al. A unified linear viscoelastic model of the cell nucleus defines the mechanical contributions of lamins and chromatin[J]. Adv Sci (Weinh), 2020, 7(8): 1901222. doi: 10.1002/advs.201901222
|
| [131] |
Sapra KT, Qin Z, Dubrovsky-Gaupp A, et al. Nonlinear mechanics of lamin filaments and the meshwork topology build an emergent nuclear lamina[J]. Nat Commun, 2020, 11(1): 6205. doi: 10.1038/s41467-020-20049-8
|
| [132] |
Khilan AA, Al-Maslamani NA, Horn HF. Cell stretchers and the LINC complex in mechanotransduction[J]. Arch Biochem Biophys, 2021, 702: 108829. doi: 10.1016/j.abb.2021.108829
|
| [133] |
|
| [134] |
Sharma P, Alsharif S, Fallatah A, et al. Intermediate filaments as effectors of cancer development and metastasis: a focus on keratins, vimentin, and nestin[J]. Cells, 2019, 8(5): 497. doi: 10.3390/cells8050497
|
| [135] |
Nakamura F, Song M, Hartwig JH, et al. Documentation and localization of force-mediated filamin A domain perturbations in moving cells[J]. Nat Commun, 2014, 5: 4656. doi: 10.1038/ncomms5656
|
| [136] |
Aragona M, Panciera T, Manfrin A, et al. A mechanical checkpoint controls multicellular growth through YAP/TAZ regulation by actin-processing factors[J]. Cell, 2013, 154(5): 1047–1059. doi: 10.1016/j.cell.2013.07.042
|
| [137] |
Nakamura F. The role of mechanotransduction in contact inhibition of locomotion and proliferation[J]. Int J Mol Sci, 2024, 25(4): 2135. doi: 10.3390/ijms25042135
|
| [138] |
Fallatah A, Anastasakis DG, Manzourolajdad A, et al. Keratin 19 binds and regulates cytoplasmic HNRNPK mRNA targets in triple-negative breast cancer[J]. BMC Mol Cell Biol, 2023, 24(1): 26. doi: 10.1186/s12860-023-00488-z
|
| [139] |
Kim SH, Kim S, Choi HI, et al. Callus formation is associated with hyperproliferation and incomplete differentiation of keratinocytes, and increased expression of adhesion molecules[J]. Br J Dermatol, 2010, 163(3): 495–501. doi: 10.1111/j.1365-2133.2010.09842.x
|
 Authors and Reviewers
Authors and Reviewers

 DownLoad:
DownLoad:

