Qianfeng Chen, Yuxia Zhong, Bohan Li, Yucong Feng, Yuandie Zhang, Tao Wei, Margaret Zaitoun, Shuang Rong, Hua Wan, Qing Feng. Acrolein-triggered atherosclerosis via AMPK/SIRT1-CLOCK/BMAL1 pathway and a protection from intermittent fasting[J]. The Journal of Biomedical Research. DOI: 10.7555/JBR.38.20240025
Citation:
Qianfeng Chen, Yuxia Zhong, Bohan Li, Yucong Feng, Yuandie Zhang, Tao Wei, Margaret Zaitoun, Shuang Rong, Hua Wan, Qing Feng. Acrolein-triggered atherosclerosis via AMPK/SIRT1-CLOCK/BMAL1 pathway and a protection from intermittent fasting[J]. The Journal of Biomedical Research. DOI: 10.7555/JBR.38.20240025
Qianfeng Chen, Yuxia Zhong, Bohan Li, Yucong Feng, Yuandie Zhang, Tao Wei, Margaret Zaitoun, Shuang Rong, Hua Wan, Qing Feng. Acrolein-triggered atherosclerosis via AMPK/SIRT1-CLOCK/BMAL1 pathway and a protection from intermittent fasting[J]. The Journal of Biomedical Research. DOI: 10.7555/JBR.38.20240025
Citation:
Qianfeng Chen, Yuxia Zhong, Bohan Li, Yucong Feng, Yuandie Zhang, Tao Wei, Margaret Zaitoun, Shuang Rong, Hua Wan, Qing Feng. Acrolein-triggered atherosclerosis via AMPK/SIRT1-CLOCK/BMAL1 pathway and a protection from intermittent fasting[J]. The Journal of Biomedical Research. DOI: 10.7555/JBR.38.20240025
Unproofed Manuscript: The manuscript has been professionally copyedited and typeset to confirm the JBR’s formatting, but still needs proofreading by the corresponding author to ensure accuracy and correct any potential errors introduced during the editing process. It will be replaced by the online publication version.
Acrolein-triggered atherosclerosis via AMPK/SIRT1-CLOCK/BMAL1 pathway and a protection from intermittent fasting
Department of Nutrition and Food Hygiene, Key Laboratory of Toxicology, School of Public Health, Nanjing Medical University, Nanjing, Jiangsu 211166, China
2.
Department of nosocomial infection management, Suzhou Wujiang District Children's Hospital, Suzhou, Jiangsu 215299, China
3.
Department of Preventive Medicine, Department of Nutritional Health and Toxicology, School of Public Health, Wuhan University of Science and Technology, Wuhan, Hubei 430065, China
4.
Healthcare center, Sir Run Run Hospital, Nanjing Medical University, Nanjing, Jiangsu 211112, China
Hua Wan, Healthcare center, Sir Run Run Hospital, Nanjing Medical University, 109 Longmian Avenue, Nanjing, Jiangsu 211112, China. E-mail: wanhua2006@njmu.edu.cn
Qing Feng, Department of Nutrition and Food Hygiene, Key Laboratory of Toxicology, School of Public Health, Nanjing Medical University, 101 Longmian Avenue, Nanjing, Jiangsu 211166, China. E-mail: qingfeng@njmu.edu.cn
Circadian clock plays a vital role in the pathological progression of cardiovascular disease (CVD). Our previous studies showed that acrolein, an environmental pollutant, promoted atherosclerosis by reducing CLOCK/BMAL1 and disturbing circadian rhythm. Whereas, intermittent fasting (IF), a diet pattern, was able to ameliorate acrolein-induced atherosclerosis. In vivo, mice were fed acrolein 3 mg/kg/day via drinking water and IF for 18h (0:00-18:00). We observed that IF decreased acrolein-accelerated the formation of aortic lesion in ApoE−/− mice. Up-regulation of NF-κB, IL-1β and TNF-α levels were found in liver and heart tissue upon acrolein exposure, while was down-regulated by IF. Interestingly, IF treatment exhibited higher AMPK, p-AMPK and SIRT1and lower MAPK expression which was caused by acrolein. Besides, circadian genes Clock/ Bmal1 expression were suppressed and disturbed treated with acrolein, while were reversed by IF. Furthermore, consistent with that in vivo, short-term starvation as a fasting cell model in vitro could improve the disorders of CLOCK/BMAL1 and raised SIRT1 via regulating AMPK, as well as ROS-MAPK induced by acrolein. In conclusion, we demonstrated that IF repressed ROS-MAPK while activated AMPK to elevate the expression of circadian clock genes to ameliorate acrolein-induced atherogenesis, which shed a novel light to prevent cardiovascular diseases.
Cardiovascular disease (CVD) remains the leading cause of death, and atherosclerosis (As) is one of the pathological bases of CVD. A growing number of studies indicate that environmental pollution shows considerable effects on development of CVD[1–2]. Acrolein, an α, β-unsaturated aldehyde is commonly found in cigarette smoke, car exhaust, cooking fumes and food. It is reported to correlate with increased risk of CVD[3]. Studies have shown that acrolein can initiate and accelerate the formation of atherosclerotic lesions through various mechanisms, such as oxidative stress, enhancement of inflammatory processes, and activation of matrix metalloproteinases[4]. In addition, acrolein exposure is thought to induce systemic dyslipidemia, an important risk factor for the development of atherosclerosis. Also, acrolein increased platelet activation and stimulation of the coagulation cascade, leading to thrombosis[5]. The prevalence of vascular disease may be decreased, when reducing acrolein exposure from known sources. Fortunately, dietary factors like olive extract,asparagus extract and dietary polyphenols could attenuated CVD caused by acrolein[6–8], which suggests that diet could provide protective effect on cardiovascular diseases resulted from environmental pollution.
Intermittent fasting (IF) refers to a diet that alternates between fasting and free eating for a period of time to prevent and treat disease, such as obesity, type 2 diabetes mellitus and CVD, which includes time-restricted feeding (TRF, limiting eating to specific periods of time), 5∶2 diet (normal diet with 5 days and fasting diet with 2 days), and alternate-day fasting (ADF, eating normally one day and fasting lightly every other day)[9]. As a dietary pattern, IF is not a direct restriction of energy intake, but a fasting during a specific time and a free diet at other times, which is safer than the energy restriction[10]. It has been demonstrated that IF can reduce risk factors associated with atherosclerosis, like inflammatory factors and blood lipid levels[11]. Moreover, IF significantly decreased adhesion of monocyte and hypercholesterolemia to ameliorate atherosclerosis in LDLR−/− mice[12]. However, the effect of IF on atherosclerosis and its potential mechanism have not been determined.
Disturbances in circadian rhythms are thought to contribute to the development of CVD[13]. Studies have demonstrated that people with chronic exposure to shift work, night work, jet lag and sleep disorders have substantially more at risk for CVD[14–15]. The circadian rhythm is based on molecular clock genes involved the core clock genes CLOCK and BMAL1. CLOCK and BMAL1 regulate the genes expression of Period (Per1, Per2) and Cytochrome (Cry1, Cry2) by a transcription-translation negative feedback loop to generate molecular oscillations[16]. Deletion or mutation of circadian clock genes leads to disturbance of biological rhythms, inflammation and metabolic disease[17]. It has been reported that knockdown of Bmal1 enhanced NF-κB signaling, increased oxidative stress and inflammatory response in mice, which implied the effect of circadian rhythms on atherosclerosis[18]. Our previous studies showed that acrolein caused disruption of circadian clock and activated MAPK (mitogen activated protein kinase) related molecules[6,19]. Now, it is still unclear whether intermittent fasting reversed circadian rhythm disturbance and higher expression of MAPK by acrolein.
IF is closely related to energy-response molecules e.g. AMPK (AMP-activated protein kinase) and SIRT1 (sirtuin1). As an energy regulator, AMPK contributes to dysregulation in diabetes, cardiometabolic diseases and cancer[20]. Activation of AMPK upregulates NAMPT (Nicotinamide Phosphoribosyl transferase), which increases the accumulation level of molecule NAD+, and drives the rhythmic activity of SIRT1. SIRT1 is a sensing molecule of nutritional status and circadian rhythm regulation, affecting the circadian clock in the brain and peripheral tissues[21]. The recent studies have confirmed that environmental pollutants induced oxidative stress, which caused disruption of circadian clock[22–23]. In addition, MAPK was able to accelerate atherosclerosis by activating ROS[24]. The crystal structure of the protein illustrated that: AMPK can phosphorylate serine sites on Per to affect the stability of Cry, while MAPK can phosphorylate the upper phase of Cry, which affects Cry-dependent transcriptional inhibition of CLOCK/BMAL1[22]. Thus, we hypothesized that AMPK-SIRT1 and ROS-MAPK participated in the regulation of circadian clock through redox signaling pathway.
Since IF actually optimizes feeding times and circadian rhythms in the various organs of mammals[25], whether IF ameliorates atherosclerosis associated with AMPK, MAPK and circadian clock genes remains uncertain. Therefore, we aimed to explore the effect of IF on acrolein-induced atherosclerosis and its potential molecular mechanism. We expect this study would provide a new perspective of IF for the prevention and treatment of cardiovascular diseases.
Materials and methods
Cell culture
HUVEC (Human Umbilical Vein Endothelial Cells), MOVAS (Murine Aortic Vascular Smooth Muscle Cells) and Raw264.7 cells were obtained from the Chinese Academy of Sciences Committee on Type Culture Collection Cell Bank (Shanghai, China). Cells were cultured in 90% Dulbecco’s-modified Eagle’s medium (DMEM, Procell, Wuhan, Hubei, China) supplemented with 10% fetal bovine serum (HAKATA, Shanghai, China), 100 U/mL penicillin and 100 mg/mL streptomycin (Beyotime, Shanghai, China) at 37 ℃with 5% CO2 incubator.
STS (Short-term starvation) was supplemented with 0.5 g/L glucose and 1% FBS in DMEM sugar-free medium (Procell, Wuhan, Hubei, China)[26]. Since it was established by the investigators based on a FMD of 1-2 days per week (energy reduction to 20-25% on fasting days), we used STS as a cellular model for IF. Cells were cultured in DMEM high sugar complete medium in Petri dishes for 2-3 days, and then switched to the STS solution described above and continued to culture for 16-24 h.
Western blotting
Cells or tissues were lysed with lysis solution (RIPA: PMSF=100:1) prepared with cold protein lysis buffer (RIPA, Beyotime, Shanghai, China) and benzyl alcohol sulfonyl fluoride (PMSF, Beyotime, Shanghai, China). The protein concentrations were quantified with the BCA protein assay (Beyotime, Shanghai, China). The protein samples were then separated on 8% SDS polyacrylamide gels and transferred to polyvinylidene difluoride (PVDF, Millpore, Boston, Massachusetts, USA) membranes. After sealing the membrane with 5% fat free milk, the primary antibody was incubated overnight at 4 °C and the secondary antibody was incubated at indoor temperature for 1h. The results were evaluated by a chemiluminescence image analysis system (Tanon, Shanghai, China).
The antibodies against used in this study were as followed: AMPKα1 (1∶1000, Cat. #380431, ZENBIO, Chengdu, Sichuan, China), p-AMPKα1 (1∶1000, Cat. #310044, ZENBIO, Chengdu, Sichuan, China), SIRT1 (1∶1000, Cat. #R25721, ZENBIO, Chengdu, Sichuan, China), p-P38 (1∶1000, Cat. #310068, ZENBIO, Chengdu, Sichuan, China), ERK (1∶1000, Cat. #R22685, ZENBIO, Chengdu, Sichuan, China), p-ERK (1∶1000, Cat. #343830, ZENBIO, Chengdu, Sichuan, China), JNK (1∶1000, Cat. #R22866, ZENBIO, Chengdu, Sichuan, China), p-JNK (1∶1000, Cat. #340810, ZENBIO, Chengdu, Sichuan, China), CLOCK (1∶1000, Cat. #R381971, ZENBIO, Chengdu, Sichuan, China), BMAL1 (1∶1000, Cat. #220406, ZENBIO, Chengdu, Sichuan, China), NF-κB (1∶1000, Cat. #222013, ZENBIO, Chengdu, Sichuan, China), IL-6 (1∶1000, Cat. #500286, ZENBIO, Chengdu, Sichuan, China), IL-1β (1∶1000, Cat. #660092, ZENBIO, Chengdu, Sichuan, China) and TNF-α (1∶1000, Cat. #346654, ZENBIO, Chengdu, Sichuan, China), P38 (1∶1000, Cat. #R27123, ZENBIO, Chengdu, Sichuan, China) and GAPDH (1∶1000, Cat. #250133, ZENBIO, Chengdu, Sichuan, China). Secondary antibodies were HRP-conjugated AffiniPure Goat Anti-Rabbit IgG and HRP-conjugated AffiniPure Goat Anti- Mouse IgG (1∶2000, Cat. #S0001 and #S0002, Affinity, Alum Creek, WV, USA). The protein expression was calculated by ImageJ software (National Institutes of Health, Bethesda, MD, USA).
RNA isolation and real-time reverse transcription PCR (qRT-PCR)
Total RNA was extracted from the samples by RNAiso Plus kit (YIFEIXUE Bio, Nanjing, Jiangsu, China) following the manufacturer’s instructions, and reverse transcription was Performed with Prime ScriptTM RT Master Mix (Takara, Dalian, Liaoning, China). Quantitative polymerase chain reaction was performed using SYBR Premix Ex Taq II (YIFEIXUE, Bio, Nanjing, Jiangsu, China) and a Roche Light Cycler 96 or 480 Real Time PCR system (Roche, Basel, Switzerland). The PCR primer sequences in this study are listed in Table 1. The relative expression levels were calculated by the 2−△△Ct method using GAPDH as the reference gene.
Table
1.
Primer sequences used for real-time reverse transcription PCR
DCFH-DA (Beyotime, Shanghai, China) was diluted as 1∶1000 in serum-free medium. Cells were collected and incubated with DCFH-DA for 20 min at 37 ℃. The cells were washed three times with serum-free medium and then twice with phosphate buffered saline (PBS). Finally, resuspend in 400 μL PBS and analyzed by flow cytometry.
siRNA and plasmid transfection
The siRNAs of AMPKα1 or the corresponding control (RiboBio, Guangzhou, Guangdong, China), AMPKα1 plasmid, or the control plasmid were transfected into cells with Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s instructions. The sequence of siRNAs of AMPKα1 was shown in Table 2.
Table
2.
Target sequence of siRNA for transfection
Twenty male ApoE−/− mice (6-7 weeks, 20–25 g) were purchased from the Model Animal Research Center of Nanjing University (Nanjing, Jiangsu, China). Mice were fed with high-fat diet (18.9% protein, 44.6% carbohydrate and36.5% kcal/g fat) for 8 weeks in the Laboratory Animal Center of Nanjing Medical University to constructed atherosclerosis models, then divided into four groups and fed under a 12h light/12h dark schedule for 8 weeks: The control group was fed freely without acrolein (Sinopharm Chemical Reagent Company, Nanjing, Jiangsu, China), the time-restricted fasting (TRF) group was fed for 6 h a day (18:00 to 24:00), the acrolein (ACR) group was fed with 3 mg/kg acrolein in water every day, and the acrolein combined with time-restricted fasting (ACR+TRF) group was fed for 6 h (18:00 to 24:00) and 3 mg/kg acrolein in drinking water every day. The mice were sacrificed to collect aorta, liver, and heart tissues.
A further sixty-four male C57BL/6J mice (6-7 weeks, 20-25 g) were purchased from Nanjing Medical University Animal Core (Nanjing, Jiangsu, China). The C57BL/6J mice were fed with a high-fat diet for 8 weeks, then divided into the same groups and treated for 12 weeks. Finally, they were sacrificed at 6 am (zeitgeber time [ZT] 0/24), 12 am (ZT6), 6 pm (ZT12), and 12 pm (ZT18) to collect plasma, aorta, liver, and heart tissues.
Animal experiments were performed in accordance with the guide for the care and use of laboratory animals, and approved by the Animal Ethics and Welfare Committee (AEWC) of Nanjing Medical University.
Oil red O staining and HE staining
Formalin-fixed aortas: The aorta was dissected, part of the aorta was fixed in 4% paraformaldehyde for 48 h, and then 4 μm sections were prepared by whole oil red O staining and paraffin embedding. Stained with the oil red O: The aorta slice fixed with 4% paraformaldehyde was taken out and placed into the configured oil red O working liquid. After staining, the slice was taken out and differentiated into 60% isopropyl alcohol until the plaque was orange and the rest of the normal aorta tissue was milky white, and then photographed. Formalin-fixed and stained Raw264.7 cells with the oil red O: Raw264.7 cells were fixed in 4% paraformaldehyde for 20-30 minutes, rinsed and placed into the configured oil red O working liquid. Finally, the cells were washed off the dye and photographed.
HE staining: After dewaxing, hydration and dehydration, the paraffin sections of aortic tissue were stained by HE, then sealed and photographed under a light microscope.
Image-pro Plus software (Version 6.0; Cybernetics of Media, Bethesda, MD) was used to analyze the staining results.
Serum lipid detection
The concentrations of plasma triglycerides (TG), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C) and cholesterol (CHOL) were determined using commercial colorimetric kits (Biolabo SAS, Maizy, France) based on GPO and CHOD-PAP detection methods, coupling enzymatic reaction, and detection of reaction end products by spectrophotometry (CLARIOstar Plus microplate reader, BMG Labtech, Ortenberg, Germany).
Statistical analysis
All experiments were presented as the mean ± standard deviation of three independent experiments. Two-tailed Student's t-test for two groups, one-way analysis of variance (ANOVA) followed by the Bonferroni multiple comparison test for two or more groups and two-way followed by Dunnett's tests were applied to compare the data between two or more groups by using GraphPad Prism 7 (GraphPad Software, La Jolla, CA, USA). P < 0.05 was considered statistically significant.
Results
IF attenuated acrolein-triggered aortic plaque area and inflammatory factors in ApoE−/− mice
To explore whether IF attenuated acrolein-triggered development of atherosclerosis, we divided the ApoE−/− mice into 4 groups, and fed them with high-fat diet (Fig. 1A). We found that body weight of mice was lower in TRF and TRF+ACR groups than that in control and ACR groups (Fig. 1B). As was shown in Supplementary Fig. 1A and 1B (available online), ACR significantly increased serum CHOL and LDL-C, whereas ACR plus TRF treatment was able to reverse this upward trend. Besides, TRF elevated HDL-C, while ACR plus TRF decreased it. In the group of acrolein treatment, the plaque area in oil red O staining of aorta significantly increased compared with the control group (Fig. 1C). The evidence has been confirmed that inflammation participates in the development of atherosclerosis[27], therefore, we detected the inflammatory factors and found NF-κB and IL-1β were downregulated in liver and heart tissue significantly in mice fed with TRF. In addition, both of them were lower with ACR plus TRF treatment than ACR alone (Fig. 1D and 1E). For ACR treatment, only IL-1β in the liver and TNF-α in the heart were apparently increased compared with the control group (Fig. 1E and 1G). Also, there was a trend in mRNA of IL-6 changes, but no statistical difference. Taken together, our findings indicated that IF may inhibit acrolein-accelerated aortic plaque and inflammation.
Figure
1.
Effects of ACR or/and TRF on atherosclerosis in ApoE−/− mice.
A: A total of 20 ApoE−/− mice with 6-7-week-old were fed with high-fat diet for 8 weeks to build a model of atherosclerosis, Then they were divided into four groups (n = 5): control group (Control), time-restricted feeding group (TRF): fed for 6 h per day (18:00 to 24:00), acrolein group (ACR): 3 mg/ (kg·day) by drinking water, and combined acrolein with time-restricted fasting group (ACR+TRF). All mice were fed with high-fat diet for another 8 weeks. B: Body weight of 4 groups of mice from 10-16weeks. ACR group and ACR+TRF group respectively compared with Control group. (n = 5). C: The aorta of ApoE−/−mice were stained with oil red O (n = 3). Representative images of aorta of ApoE−/−mice were observed by microscope and percentage of stain area after treatment with ACR or/and TRF for 8 weeks. D: Quantitative reverse transcription-PCR detection of mRNA expression of NF-κB in the heart and liver of the mice (n = 3)). E: Quantitative reverse transcription-PCR detection of mRNA expression of IL-1β in the heart and liver of ApoE−/− mice (n = 3)). F: Quantitative reverse transcription-PCR detection of mRNA expression of IL-6 in the heart and liver of ApoE−/− mice (n = 3)). G: Quantitative reverse transcription-PCR detection of mRNA expression of TNF-α in the heart and liver of ApoE−/− mice (n = 3)). Data were expressed as mean ± standard deviation of at least three experiments and analyzed by one-way analysis of variance (ANOVA), followed by Bonferroni multiple comparison test except that statistical analysis for B was performed by two-way ANOVA followed by Dunnett's tests. ns: no significance, *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001. Abbreviations: ApoE−/−, ApoE-deficient; NF-κB, nuclear factor kappa-B, IL-1β, Interleukin-1β; IL-6, Interleukin 6; TNF-α, tumor necrosis factor; Gapdh, glyceraldehyde 3-phosphate dehydrogenase.
TRF may enhance the expression of AMPK, SIRT1 but inhibit MAPK caused by ACR in ApoE−/− mice.
It has been shown that the liver is the largest energy metabolizing organ in the human body[28]. The energy response molecules are usually activated by hungry or fasting[29]. Therefore, we measured energy-related proteins after the ApoE−/− mice were given the indicated treatment. We found that the protein of AMPK, p-AMPK and SIRT1 in the liver were inhibited with ACR treatment but enhanced by TRF compared with the control group (Fig. 2A–2D), though there is no significance. Moreover, comparing ACR+TRF group with ACR group, these energy indicators could be reversed in an upward trend. Meanwhile, the same trend was observed after ACR and/or TRF treatment in both liver and heart tissue in mRNA level (Fig. 2E–2H). These results displayed that IF may ameliorate the effects of acrolein on energy response molecules AMPK-SIRT1.
Figure
2.
Expression of AMPK, SIRT1 and MAPK after the indicated treatment in ApoE−/− mice.
The ApoE−/− mice were treated with ACR or/and TRF for 8 weeks. A: The protein levels of AMPK, p-AMPK and SIRT1 were detected by western blotting in the liver of ApoE−/−mice (n = 3). B: The semi-quantitative analyses of AMPK in A (n = 3), using GAPDH as a reference. C: The semi-quantitative analyses of p-AMPK in A (n = 3), using GAPDH as a reference. D: The semi-quantitative analyses of SIRT1 in A (n = 3), using GAPDH as a reference. E: Quantitative reverse transcription-PCR detection of mRNA expression of Ampk in the liver of ApoE−/−mice (n = 3). F: Quantitative reverse transcription-PCR detection of mRNA expression of Ampk in the heart of ApoE−/−mice (n = 3). G: Quantitative reverse transcription-PCR detection of mRNA expression of Sirt1 in the liver of ApoE−/−mice (n = 3). H: Quantitative reverse transcription-PCR detection of mRNA expression of Sirt1 in the heart of ApoE−/−mice (n = 3). I: Western blotting detection of protein expression of JNK, p-JNK, ERK, p-ERK and p-P38 in the liver of ApoE−/−mice (n = 3). J: The semi-quantitative analyses of JNK in H, using P38 as a reference (n = 3). K: The semi-quantitative analyses of p-JNK in H, using P38 as a reference (n = 3). L: The semi-quantitative analyses of ERK in H, using P38 as a reference (n = 3). M: The semi-quantitative analyses of p-P38 in H, using P38 as a reference (n = 3). and Data were expressed as mean ± standard deviation of at least three experiments and analyzed by one-way ANOVA, followed by the Bonferroni multiple comparison test. ns: no significance, *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001. Abbreviations: ApoE−/−, ApoE-deficient; AMPK, AMP-activated protein kinase; p-AMPK, phospho-AMP-activated protein kinase; SIRT1, sirtuin 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; MAPK, mitogen-activated protein kinases; JNK, c-Jun N-terminal kinase; p-JNK, phosphorylated c-Jun N-terminal kinase; ERK, extracellular regulated protein kinases; p-ERK, phosphorylated extracellular regulated protein kinases; P38, P38 mitogen-activated protein kinase; p-P38, phosphorylated P38 mitogen-activated protein kinase.
Our previous studies have verified that acrolein irritated MAPK expression[6,19]. To explore further, the protein expression of JNK, p-JNK, ERK, p-ERK and p-P38 decreased obviously after TRF, but did not rise significantly upon acrolein compared with control group in protein level, except ERK (Fig. 2H and 2L). As was shown in Fig. 2J–2M, MAPK was down-regulated apparently comparing the ACR + TRF group with ACR group. These results revealed that IF could block acrolein-promoted the expression of MAPK.
TRF alleviated circadian rhythm disturbances and elevated Clock and Bmal1 upon acrolein in the mice
As the liver is also the largest peripheral organ of the circadian clock[30], we examined circadian clock genes expression of heart and liver tissue in ApoE−/− mice. As shown in Fig. 3A–2C, ACR exposure resulted in CLOCK and BMAL1 significantly decrease in liver. Besides, Bmal1 expression in heart and liver was up-regulated once ApoE−/− mice were treated with TRF (Fig. 3F and 3G). However, Clock level elevated only in the liver (Fig. 3D). Comparing ACR+TRF group with ACR group, we observed that Clock and Bmal1 could be reversed by TRF, showing an upward trend in the liver or heart (Fig. 3D and 3G). Taken together, the reversal effect of ACR by intermittent fasting may be related to intermittent fasting improving circadian clock.
Figure
3.
Expression of CLOCK and BMAL1 with treated ACR or/and TRF in the mice.
The ApoE−/− mice were treated with ACR or/and TRF for 8 weeks. A: The protein levels of CLOCK and BMAL1 were detected by western blotting in liver of ApoE−/− mice (n = 3). B: The semi-quantitative analyses of CLOCK in A (n = 3), using GAPDH as a reference. C: The semi-quantitative analyses of BMAL1 in A (n = 3), using GAPDH as a reference. D: The mRNA expression of Clock was detected by quantitative reverse transcription-PCR in the liver of ApoE−/− mice (n = 3). E: The mRNA expression of Clock was detected by quantitative reverse transcription-PCR in the heart of ApoE−/− mice (n = 3). F: The mRNA expression of Bmal1 was detected by quantitative reverse transcription-PCR in the liver of ApoE−/− mice (n = 3). G: The mRNA expression of Bmal1 was detected by quantitative reverse transcription-PCR in the heart of ApoE−/− mice (n = 3). H: The mRNA expression of Clock was detected by quantitative reverse transcription-PCR at 6 am (ZT0), 12 am (ZT6), 6 pm (ZT12) and 12 pm (ZT18) in liver of C57BL /6J mice (n = 3). TRF group and ACR group respectively compared with Control group. . I: The mRNA expression of Bmal1 was detected by quantitative reverse transcription-PCR at 6 am (ZT0), 12 am (ZT6), 6 pm (ZT12) and 12 pm (ZT18) in liver of C57BL /6J mice (n = 3). TRF group compared with Control group. J: The mRNA expression of Clock at 6 am (ZT0), 12 am (ZT6), 6 pm (ZT12) and 12 pm (ZT18) time in heart of C57BL /6J mice (n = 3). TRF group and ACR group respectively compared with Control group.. K: The mRNA expression of Bmal1 at 6 am (ZT0), 12 am (ZT6), 6 pm (ZT12) and 12 pm (ZT18) time in heart of C57BL /6J mice (n = 3). TRF group and ACR+TRF group respectively compared with Control group.. Data were expressed as mean ± standard deviation of at least three experiments and analyzed by ANOVA, followed by the Bonferroni multiple comparison test. ns: no significance, *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001. Abbreviations: ApoE−/−, ApoE-deficient; Gapdh, glyceraldehyde 3-phosphate dehydrogenase; Bmal1, brain and muscle ARNT-like 1; Clock, circadian locomotor output cycles kaput.
"Zeitgeber" can refer to any environmental cue that synchronizes 24-hour rhythmic behavior. To investigate the effect of intermittent fasting on circadian rhythm, we measured the rhythm of biogenic genes in C57B6J/L mice. From Fig. 3H–3K, the lower expression of Clock and Bmal1 were found in the heart and liver during the day (ZT6 or ZT12), then they were alleviated and reached at higher level with the arrival of night at ZT18 or ZT24 (0). The rhythmicity of the TRF group was consistent with that of the control group, showing a significantly increase of Clock and Bmal1 at night. However, the expression of Clock and Bmal1 rhythmically disappeared in ACR group. Furthermore, TRF was able to restore the acrolein-induced lower expression level of Clock and Bmal1 (Fig. 3H–3K). These results demonstrated that IF could modify acrolein-induced circadian rhythm disturbances.
Short-term starvation (STS) decreased acrolein-stimulated cell foam and ROS-MAPK
To further explore the mechanism of TRF on atherosclerosis triggered by acrolein, we used short-term starvation (STS) as a cell model in simulating IF (seen in 2.1). ACR exposure enhanced the uptake of cell lipid droplets in macrophage, while were decreased by STS treatment (Fig. 4A and 4B). The results verified that STS reduced macrophage foam.
Figure
4.
Effects of ACR or/and STS on cell lipid droplet uptake, ROS and MAPK expression in cells.
A: Raw264.7 cells were treated with ACR (10μmol/L) or/and STS and ox-LDL (50μg/mL) for 24 h. The cells were stained with oil red O and photographed under a microscope. Scale bars, 100 μm. B: The semi-quantitative analyses of lipid droplet uptake in A. C: ROS detection by flowcytometry in HUVEC and MOVAS with ACR or/and STS treatment for 24 h. (D) The semi-quantitative analyses of ROS in C. (E) The protein expression of JNK, p-JNK, ERK, p-ERK and p-P38 was determined by western blotting with ACR (0, 5, 10, 20 μmol/L) exposure for 24 h in MOVAS, using P38 as a reference. (F) The protein expression of JNK, p-JNK, ERK, p-ERK and p-P38 was determined by western blotting with ACR (10 μmol/L) or/and STS exposure for 24 h, using P38 as a reference. Data were expressed as mean ± standard deviation of at least three experiments and analyzed by one-way ANOVA, followed by the Bonferroni multiple comparison test. ns: no significance, *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001 (n = 3). Abbreviations: ox-LDL: oxidized low-density lipoprotein; ROS: reactive oxygen species; MAPK, mitogen-activated protein kinases; JNK, c-Jun N-terminal kinase; p-JNK, phosphorylated c-Jun N-terminal kinase; ERK, extracellular regulated protein kinases; p-ERK, phosphorylated extracellular regulated protein kinases; P38, P38 mitogen-activated protein kinase; p-P38, phosphorylated P38 mitogen-activated protein kinase.
As ROS-MAPK is a vital mechanism of atherogenesis, we measured ROS in endothelial and smooth muscle cells. As was seen in Fig. 4C and 4D, STS alone caused an elevation of ROS slightly, while ACR remarkably increased ROS. Whereas, the elevation was inhibited with STS presence. Interestingly, the protein expression of JNK, p-JNK, ERK, p-ERK, and p-P38 was raised when cells were treated with various doses of ACR (Fig. 4E, Supplementary Fig. 2A–2E [available online]). Conversely, the protein expression of MAPK-related molecules was alleviated by STS treatment (Fig. 4F, Supplementary Fig. 2F–2J). All these indicated that STS could inhibit ROS and MAPK stimulated by ACR.
STS promoted acrolein-induced lower expression of AMPK, SIRT1 and CLOCK-BMAL1 in HUVEC and MOVAS
The mRNA and protein level of AMPK, p-AMPK and SIRT1 decreased when HUVEC and MOVAS were treated with the indicated dose of acrolein (Fig. 5A and 5B, Supplementary Fig. 3A and 3B [available online]), while they elevated distinctly when the cells were treated with STS (Fig. 5C, Supplementary Fig. 3C and 3D). Besides, compared to ACR group, the expression of AMPK, p-AMPK and SIRT1 were increased after ACR plus STS treatment (Fig. 5D, Supplementary Fig. 3E and 3F). Furthermore, expression of protein and mRNA of CLOCK and BMAL1reduced by various doses of ACR (Fig. 5E and 5F, Supplementary Fig. 4A and 4B [available online]). However, when STS was applied, both CLOCK and BMAL1 were up-regulated (Fig. 5G, Supplementary Fig. 4C and 4D). Compared ACR+STS with ACR group, CLOCK and BMAL1 were up-regulated (Fig. 5H, Supplementary Fig. 4E and 4F). The above results showed that IF may reversed acrolein-induced lower expression of AMPK, SIRT1 and circadian clock, thereby alleviating the mechanism of acrolein-accelerated atherosclerosis. The results are consistent with that in the animal study.
Figure
5.
Effects of acrolein or/and STS on expression of AMPK, SIRT1, CLOCK and BMAL1 in HUVEC and MOVAS.
A: The protein levels of AMPK, p-AMPK, SIRT1 were determined by western blotting with various doses of ACR (5, 10, 20 μmol/L) treatment for 24 h in HUVEC and MOVAS. B: The mRNA levels of AMPK and SIRT1 were determined by quantitative reverse transcription-PCR with various doses of ACR (5, 10, 20 μmol/L) treatment for 24 h in HUVEC and MOVAS. C: The protein levels of AMPK, p-AMPK, SIRT1 were determined by western blotting with STS treatment for 24 h in HUVEC and MOVAS. D: The protein levels of AMPK, p-AMPK, SIRT1 were determined by western blotting with ACR (10μmol/L) or/and STS treatment for 24 h in HUVEC and MOVAS. E: The protein levels of CLOCK and BMAL1 were determined by western blotting with various doses of ACR (5, 10, 20 μmol/L) treatment for 24 h in HUVEC and MOVAS. F: The mRNA levels of CLOCK and BMAL1 were determined by quantitative reverse transcription-PCR with various doses of ACR (5, 10, 20 μmol/L) treatment for 24 h in HUVEC and MOVAS. G: The protein levels of CLOCK and BMAL1 were determined by western blotting with STS treatment for 24 h in HUVEC and MOVAS. H: The protein levels of CLOCK and BMAL1 were determined by western blotting with ACR (10μmol/L) or/and STS treatment for 24 h in HUVEC and MOVAS. Data were expressed as mean ± standard deviation of at least three experiments and analyzed by one-way ANOVA, followed by the Bonferroni multiple comparison test. ns: no significance, *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001. Abbreviations: AMPK, AMP-activated protein kinase; p-AMPK, phospho-AMP-activated protein kinase; SIRT1, sirtuin 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; BMAL1, brain and muscle ARNT-like 1; CLOCK, circadian locomotor output cycles kaput.
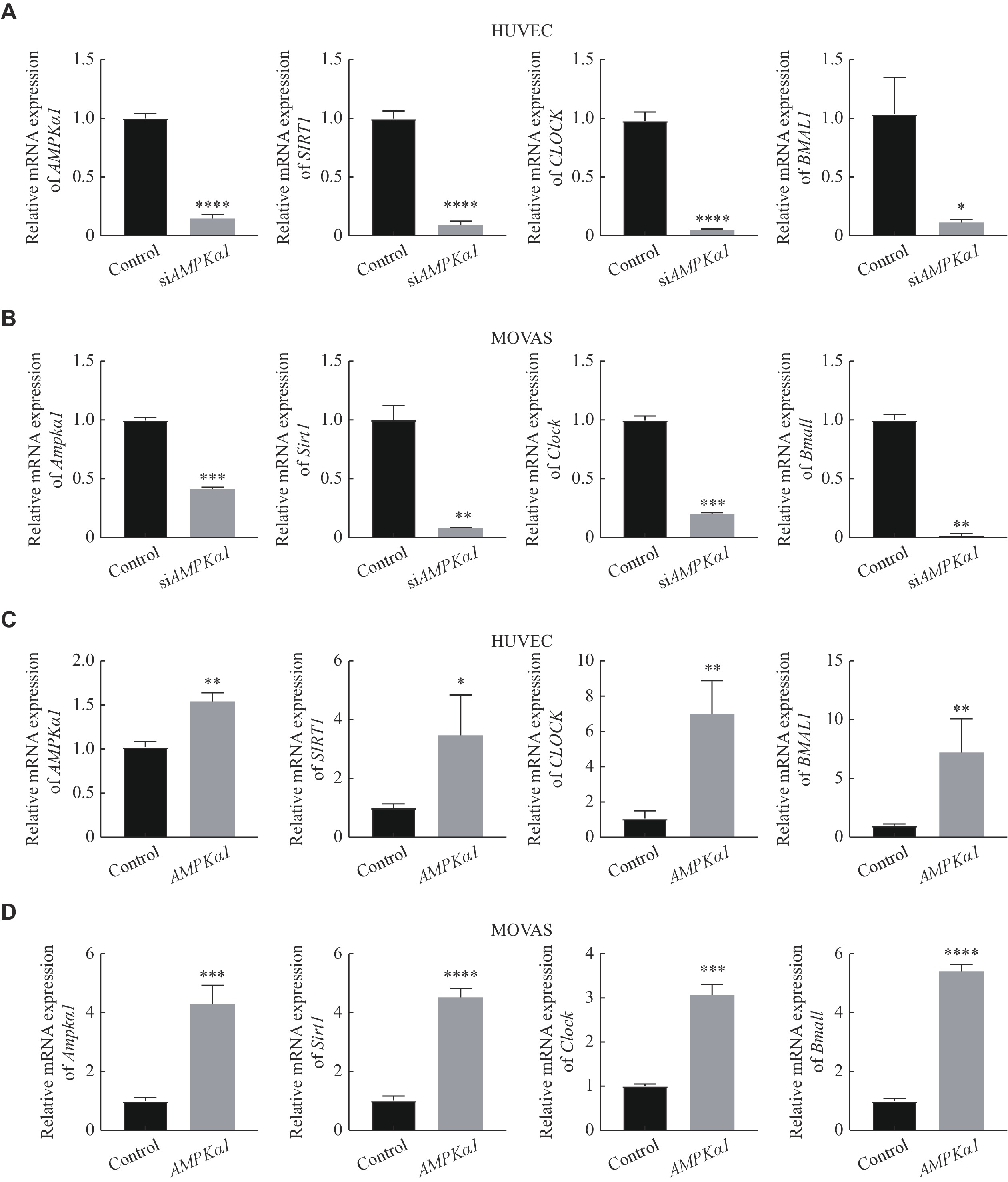
AMPK regulated SIRT1 and CLOCK/BMAL1 in HUVEC and MOVAS
We further explored whether AMPK regulated the expression of SIRT1 and CLOCK/BMAL1 in HUVEC and MOVAS. As is shown in Fig. 6A and 6B, we observed that SIRT1 and CLOCK/BMAL1 significantly decreased in mRNA level by knocking down of AMPKα1. Whereas, the expression of SIRT1, CLOCK and BMAL1 were remarkably up-regulated by overexpression of AMPKα1 (Fig. 6C and 6D). Therefore, we verified that AMPK was able to directly regulate the expression of SIRT1 and CLOCK/BMAL1.
Figure
6.
regulated CLOCK and BMAL1 in HUVEC and MOVAS.
A: The mRNA levels of AMPK, SIRT1, CLOCK and BMAL1 were detected by quantitative reverse transcription-PCR with siAMPKα1 cultured for 24 h in HUVEC. B: The mRNA levels of Ampk, Sirt1, Clock and Bmal1 were detected by quantitative reverse transcription-PCR with siAMPKα1 cultured for 24 h in MOVAS. C: The mRNA levels of AMPK, SIRT1, CLOCK and BMAL1 were detected by quantitative reverse transcription-PCR when HUVEC were transfected with AMPKα1 plasmid and cultured for 24 h. D: The mRNA levels of Ampk, Sirt1, Clock and Bmal1 were detected by quantitative reverse transcription-PCR when MOVAS were transfected with AMPKα1 plasmid and cultured for 24 h. Data were expressed as mean ± standard deviation of at least three experiments and analyzed by two-tailed Student's t-test. ns: no significance, *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001. Abbreviations: siAMPKα1∶ AMPKα1-small interfering RNA.; AMPK, AMP-activated protein kinase; p-AMPK, phospho-AMP-activated protein kinase; SIRT1, sirtuin 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; BMAL1, brain and muscle ARNT-like 1; CLOCK, circadian locomotor output cycles kaput.
Recently, IF has been shown to act as a dietary pattern, regulating circadian clock to make feeding time and the various organs of mammals to reach the optimal circadian rhythm, which reduces the risk of cardiovascular disease[31]. Our previous studies confirmed that acrolein reduced CLOCK/BMAL1 and triggered circadian rhythm disturbance, while dietary factor was able to ameliorate them[6,19]. In the present study, we displayed that IF repressed acrolein-accelerated inflammation and atherosclerotic plaque. Moreover, IF elevated the reduction of AMPK, SIRT1, CLOCK and BMAL1, as well as restored circadian rhythm disorder by ACR exposure in mice. Consistent with vivo experiment, STS treatment decreased MAPK in MOVAS. Also, STS inhibited acrolein-stimulated ROS and cell foam. To our knowledge, this is the first report of IF as an intervention modality for pollution-triggered atherosclerosis through AMPK-SIRT1/ROS-MAPK/ CLOCK-BMAL1 pathway, offering new aspects into mechanisms of atherosclerosis.
Acrolein triggers and accelerates atherosclerosis by increasing LDL-C induced macrophages foam formation, stimulating ROS production[32–34], releasing inflammatory factors like IL-1β, IL-6 and TNF-α in HUVEC cells[35], as well as directly exacerbating the formation of atherosclerotic lesions in the aortic valve and aortic arch[32,36]. Consistent with the previous reports[6,19], the current results indicated that ACR suppressed the expression of circadian clock genes, accelerated the development of atherosclerosis. In addition, we have identified that ACR inhibited energy-responsive molecules AMPK-SIRT1 in accelerating atherogenesis.
AMPK is involved in anti-atherogenic effects, such as affecting inflammatory factors and reducing cell foam. A recent study has revealed that SIRT1 could protect against inflammation and atherosclerotic plaque development[37]. Furthermore, AMPK and SIRT1 are closely associated with circadian rhythms[38–39]. And fasting activates AMPK. Our results showed that AMPK expressed in accordance with SITRT1, CLOCK and BMAL1 when AMPK was knocked down or plasmid transfected, which suggested that IF may be involved in atherosclerotic effects through AMPK regulating CLOCK and BMAL1.
MAPK plays an important role in the progression of atherosclerosis- stimulated by ACR in the current study. In addition, p38 MAPK can be involved in inflammation in different environments and cell types[40]. Several recent studies have confirmed that ROS and inflammatory factors were able to activate MAPK, which affected on regulating circadian clock genes[21,41–42]. Our findings verified that IF reduced ROS, MAPK and reversed the circadian rhythm disorder caused by ACR.
Asynchrony between circadian rhythms and metabolism remarkedly increases the risk of cardiovascular diseases[16]. In addition, IF optimized circadian rhythms and reduced the risk of metabolic disease[25]. We further explored that the rhythmicity of IF up-regulated the expression of Clock and Bmal1. It is interesting that our data showed Clock in mRNA level was lower in the heart than that in the liver and returned to initial levels during the day. The reason was probably that the liver is the largest peripheral biological clock organ[21] and Clock gene may have been influenced by the oscillation of the circadian rhythm.
The studies have reported that caloric restriction (CR), is just the reduction of energy intake without malnutrition[43], while TRF, alternative feeding methods of CR, restricts daily eating to a period ranging from 4-12 hours[44]. In our experiments, mice in TRF group were fed during 18:00-24:00 freely. Consequently, CLOCK-BMAL1 were up-regulated by TRF, not CR, which was in line with results of Jamshed H et al[45]. In vitro, STS of 24h would not cause severe physiological implication, while may protect normal fibroblasts by regulating cell cycle[46]. Consistent with our findings, in the early stage of atherosclerosis, IF could reduce aortic plaque[47] and increased HDL-C[48]. However, IL-6 expression of mice treated with TRF raised and a recent study have demonstrated that IF may increase inflammatory factors in LDL−/− mice[49]. In the intermediate stage, STS decreased macrophage foaming, lipid accumulation and ROS. In advanced stages, IF reduced myocardial infarction and post-infarction and cardiomyocyte hypertrophy, thereby promoting cardiac remodeling and survival[50].
In conclusion, our study identified that IF inhibited acrolein-induced ROS-MAPK while activated AMPK and circadian clock genes expression, thereby ameliorated atherogenesis. Hence, IF may be a new approach against environmental pollution.
However, there are still shortcomings in this study. In animal study, owing to aggressive behaviors, one of mice died during the experiment, so we uniformly chose five mice as a group. About experimental indicators, we focused on the development of early atherosclerosis in mice, such as the area of aortic plaques and levels of inflammation, and did not perform late plaque stability measurements and the ratio of fibroblasts to collagen content. In addition, due to the technical problems, we did not extract mouse aortic progenitor cells and failed to fully characterize the biological properties of atherosclerotic mice in terms of cells. Taken together, the AMPK-SIRT1/ROS-MAPK/CLOCK-BMAL1 regulatory axis may become a new target for atherosclerosis prevention and treatment in the future, once confirmed by other investigators.
Fundings
The present study is supported by the Priority Academic Program Development of Jiangsu Higher Education Institutions and the Key Project of Jiangsu Commission of Health (Grant No. ZD2022012).
Acknowledgments:
We acknowledge and appreciate our professor and institutional colleagues for their experimental technical support.
Kaufman JD, Adar SD, Barr RG, et al. Association between air pollution and coronary artery calcification within six metropolitan areas in the USA (the Multi-Ethnic Study of Atherosclerosis and Air Pollution): a longitudinal cohort study[J]. Lancet, 2016, 388 (10045): 696–704. doi: 10.1016/S0140-6736(16)00378-0
[2]
de Bont J, Jaganathan S, Dahlquist M, et al. Ambient air pollution and cardiovascular diseases: an umbrella review of systematic reviews and meta-analyses[J]. J Intern Med, 2022, 291 (6): 779–800. doi: 10.1111/joim.13467
[3]
DeJarnett N, Conklin DJ, Riggs DW, et al. Acrolein exposure is associated with increased cardiovascular disease risk[J]. J Am Heart Assoc, 2014, 3 (4): e000934. doi: 10.1161/JAHA.114.000934
[4]
Moghe A, Ghare S, Lamoreau B, et al. Molecular mechanisms of acrolein toxicity: relevance to human disease[J]. Toxicol Sci, 2015, 143 (2): 242–255. doi: 10.1093/toxsci/kfu233
[5]
Zirak MR, Mehri S, Karimani A, et al. Mechanisms behind the atherothrombotic effects of acrolein, a review[J]. Food Chem Toxicol, 2019, 129: 38–53. doi: 10.1016/j.fct.2019.04.034
[6]
Chen L, Wu X, Zeb F, et al. Acrolein-induced apoptosis of smooth muscle cells through NEAT1-Bmal1/Clock pathway and a protection from asparagus extract[J]. Environ Pollut, 2020, 258: 113735. doi: 10.1016/j.envpol.2019.113735
[7]
Xu Y, Wu L, Chen A, et al. Protective effects of olive leaf extract on acrolein-exacerbated myocardial infarction via an endoplasmic reticulum stress pathway[J]. Int J Mol Sci, 2018, 19 (2): 493. doi: 10.3390/ijms19020493
[8]
Zhou Y, Jin W, Wu Q, et al. Acrolein: formation, health hazards and its controlling by dietary polyphenols[J]. Crit Rev Food Sci Nutr, 2023, doi: 10.1080/10408398.2023.2214625.
[9]
Vasim I, Majeed CN, DeBoer MD. Intermittent fasting and metabolic health[J]. Nutrients, 2022, 14 (3): 631. doi: 10.3390/nu14030631
[10]
Varady KA. Intermittent versus daily calorie restriction: which diet regimen is more effective for weight loss?[J]. Obes Rev, 2011, 12 (7): e593–e601.
[11]
Malinowski B, Zalewska K, Węsierska A, et al. Intermittent fasting in cardiovascular disorders—an overview[J]. Nutrients, 2019, 11 (3): 673. doi: 10.3390/nu11030673
[12]
Chen Y, Su J, Yan Y, et al. Intermittent fasting inhibits high-fat diet-induced atherosclerosis by ameliorating hypercholesterolemia and reducing monocyte chemoattraction[J]. Front Pharmacol, 2021, 12: 719750. doi: 10.3389/fphar.2021.719750
[13]
Schilperoort M, van den Berg R, Bosmans LA, et al. Disruption of circadian rhythm by alternating light-dark cycles aggravates atherosclerosis development in APOE*3-Leiden. CETP mice[J]. J Pineal Res, 2020, 68 (1): e12614. doi: 10.1111/jpi.12614
[14]
Huang T, Mariani S, Redline S. Sleep irregularity and risk of cardiovascular events: the multi-ethnic study of atherosclerosis[J]. J Am Coll Cardiol, 2020, 75 (9): 991–999. doi: 10.1016/j.jacc.2019.12.054
[15]
Kervezee L, Shechter A, Boivin DB. Impact of shift work on the circadian timing system and health in women[J]. Sleep Med Clin, 2018, 13 (3): 295–306. doi: 10.1016/j.jsmc.2018.04.003
[16]
Crnko S, Du Pré BC, Sluijter JPG, et al. Circadian rhythms and the molecular clock in cardiovascular biology and disease[J]. Nat Rev Cardiol, 2019, 16 (7): 437–447. doi: 10.1038/s41569-019-0167-4
[17]
Sun L, Ma J, Turck CW, et al. Genome-wide circadian regulation: a unique system for computational biology[J]. Comput Struct Biotechnol J, 2020, 18: 1914–1924. doi: 10.1016/j.csbj.2020.07.002
[18]
Xie M, Tang Q, Nie J, et al. BMAL1-downregulation aggravates porphyromonas gingivalis-induced atherosclerosis by encouraging oxidative stress[J]. Circ Res, 2020, 126 (6): e15–e29.
[19]
Wu X, Chen L, Zeb F, et al. Clock-Bmal1 mediates MMP9 induction in acrolein-promoted atherosclerosis associated with gut microbiota regulation[J]. Environ Pollut, 2019, 252: 1455–1463. doi: 10.1016/j.envpol.2019.06.042
[20]
Ge Y, Zhou M, Chen C, et al. Role of AMPK mediated pathways in autophagy and aging[J]. Biochimie, 2022, 195: 100–113. doi: 10.1016/j.biochi.2021.11.008
[21]
Masri S. Sirtuin-dependent clock control: new advances in metabolism, aging and cancer[J]. Curr Opin Clin Nutr Metab Care, 2015, 18 (6): 521–527. doi: 10.1097/MCO.0000000000000219
[22]
Li H, Kilgallen AB, Münzel T, et al. Influence of mental stress and environmental toxins on circadian clocks: implications for redox regulation of the heart and cardioprotection[J]. Br J Pharmacol, 2020, 177 (23): 5393–5412. doi: 10.1111/bph.14949
[23]
Sundar IK, Sellix MT, Rahman I. Redox regulation of circadian molecular clock in chronic airway diseases[J]. Free Radic Biol Med, 2018, 119: 121–128. doi: 10.1016/j.freeradbiomed.2017.10.383
[24]
Duan L, Zhao Y, Jia J, et al. Myeloid-restricted CD68 deficiency attenuates atherosclerosis via inhibition of ROS-MAPK-apoptosis axis[J]. Biochim Biophys Acta Mol Basis Dis, 2023, 1869 (5): 166698. doi: 10.1016/j.bbadis.2023.166698
[25]
Adafer R, Messaadi W, Meddahi M, et al. Food timing, circadian rhythm and chrononutrition: a systematic review of time-restricted eating's effects on human health[J]. Nutrients, 2020, 12 (12): 3770. doi: 10.3390/nu12123770
[26]
Di Tano M, Raucci F, Vernieri C, et al. Synergistic effect of fasting-mimicking diet and vitamin C against KRAS mutated cancers[J]. Nat Commun, 2020, 11 (1): 2332. doi: 10.1038/s41467-020-16243-3
[27]
Wolf D, Ley K. Immunity and inflammation in atherosclerosis[J]. Circ Res, 2019, 124 (2): 315–327. doi: 10.1161/CIRCRESAHA.118.313591
[28]
Matsubara Y, Kiyohara H, Teratani T, et al. Organ and brain crosstalk: the liver-brain axis in gastrointestinal, liver, and pancreatic diseases[J]. Neuropharmacology, 2022, 205: 108915. doi: 10.1016/j.neuropharm.2021.108915
[29]
González A, Hall MN, Lin SC, et al. AMPK and TOR: the yin and yang of cellular nutrient sensing and growth control[J]. Cell Metab, 2020, 31 (3): 472–492. doi: 10.1016/j.cmet.2020.01.015
[30]
Pan X, Mota S, Zhang B. Circadian clock regulation on lipid metabolism and metabolic diseases[M]//Jiang X. Lipid Transfer in Lipoprotein Metabolism and Cardiovascular Disease. Singapore: Springer, 2020: 53–66.
[31]
Alfarhan M, Jafari E, Narayanan SP. Acrolein: a potential mediator of oxidative damage in diabetic retinopathy[J]. Biomolecules, 2020, 10 (11): 1579. doi: 10.3390/biom10111579
[32]
Rom O, Korach-Rechtman H, Hayek T, et al. Acrolein increases macrophage atherogenicity in association with gut microbiota remodeling in atherosclerotic mice: protective role for the polyphenol-rich pomegranate juice[J]. Arch Toxicol, 2017, 91 (4): 1709–1725. doi: 10.1007/s00204-016-1859-8
[33]
Hong J, Song Y, Xie J, et al. Acrolein promotes aging and oxidative stress via the stress response factor DAF-16/FOXO in Caenorhabditis elegans[J]. Foods, 2022, 11 (11): 1590. doi: 10.3390/foods11111590
[34]
Jiang L, Luo S, Qiu T, et al. Bidirectional role of reactive oxygen species during inflammasome activation in acrolein-induced human EAhy926 cells pyroptosis[J]. Toxicol Mech Methods, 2021, 31 (9): 680–689. doi: 10.1080/15376516.2021.1953204
[35]
Yu J, Jiang Q, Liu N, et al. Apigenin and apigenin-7, 4'-O-dioctanoate protect against acrolein-aggravated inflammation via inhibiting the activation of NLRP3 inflammasome and HMGB1/MYD88/NF-κB signaling pathway in Human umbilical vein endothelial cells (HUVEC)[J]. Food Chem Toxicol, 2022, 168: 113400. doi: 10.1016/j.fct.2022.113400
[36]
Srivastava S, Sithu SD, Vladykovskaya E, et al. Oral exposure to acrolein exacerbates atherosclerosis in apoE-null mice[J]. Atherosclerosis, 2011, 215 (2): 301–308. doi: 10.1016/j.atherosclerosis.2011.01.001
[37]
D'Onofrio N, Servillo L, Balestrieri ML. SIRT1 and SIRT6 signaling pathways in cardiovascular disease protection[J]. Antioxid Redox Signal, 2018, 28 (8): 711–732. doi: 10.1089/ars.2017.7178
[38]
Jordan SD, Lamia KA. AMPK at the crossroads of circadian clocks and metabolism[J]. Mol Cell Endocrinol, 2013, 366 (2): 163–169. doi: 10.1016/j.mce.2012.06.017
[39]
Imai SI, Guarente L. NAD+ and sirtuins in aging and disease[J]. Trends Cell Biol, 2014, 24 (8): 464–471. doi: 10.1016/j.tcb.2014.04.002
[40]
Reustle A, Torzewski M. Role of p38 MAPK in atherosclerosis and aortic valve sclerosis[J]. Int J Mol Sci, 2018, 19 (12): 3761. doi: 10.3390/ijms19123761
[41]
Goldsmith CS, Bell-Pedersen D. Diverse roles for MAPK signaling in circadian clocks[J]. Adv Genet, 2013, 84: 1–39.
[42]
Li B, Li D, Ni H, et al. The circadian clock regulator Bmal1 affects traumatic brain injury in rats through the p38 MAPK signalling pathway[J]. Brain Res Bull, 2022, 178: 17–28. doi: 10.1016/j.brainresbull.2021.11.003
[43]
Duregon E, Pomatto-Watson LCDD, Bernier M, et al. Intermittent fasting: from calories to time restriction[J]. GeroScience, 2021, 43 (3): 1083–1092. doi: 10.1007/s11357-021-00335-z
[44]
Naous E, Achkar A, Mitri J. Intermittent fasting and its effects on weight, glycemia, lipids, and blood pressure: a narrative review[J]. Nutrients, 2023, 15 (16): 3661. doi: 10.3390/nu15163661
[45]
Jamshed H, Beyl RA, Della Manna DL, et al. Early time-restricted feeding improves 24-hour glucose levels and affects markers of the circadian clock, aging, and autophagy in humans[J]. Nutrients, 2019, 11 (6): 1234. doi: 10.3390/nu11061234
[46]
Pignatta S, Cortesi M, Arienti C, et al. Effects of radiotherapy and short-term starvation combination on metastatic and non-tumor cell lines[J]. DNA Repair (Amst), 2020, 95: 102949. doi: 10.1016/j.dnarep.2020.102949
[47]
Chen Y, Su J, Yan Y, et al. Intermittent fasting inhibits high-fat diet-induced atherosclerosis by ameliorating hypercholesterolemia and reducing monocyte chemoattraction[J]. Front Pharmacol, 2021, 12: 719750.
[48]
Kavanagh K, Bashore AC, Davis MA, et al. Early time-restricted feeding improves high-density lipoprotein amount and function in nonhuman primates, without effects on body composition[J]. Obesity (Silver Spring), 2023, 31 Suppl 1 (Suppl 1): 75–84.
[49]
Dorighello GG, Rovani JC, Luhman CJF, et al. Food restriction by intermittent fasting induces diabetes and obesity and aggravates spontaneous atherosclerosis development in hypercholesterolaemic mice[J]. Br J Nutr, 2014, 111 (6): 979–986. doi: 10.1017/S0007114513003383
[50]
Okoshi K, Cezar MDM, Polin MAM, et al. Influence of intermittent fasting on myocardial infarction-induced cardiac remodeling[J]. BMC Cardiovasc Disord, 2019, 19 (1): 126. doi: 10.1186/s12872-019-1113-4
 Authors and Reviewers
Authors and Reviewers

 DownLoad:
DownLoad: