Lee Sang R., Yang Hyun, Jo Seong Lae, Lee Young Ho, Lee Hye Won, Park Bae-keun, Hong Eui-Ju. Suppressed estrogen supply via extra-ovarian progesterone receptor membrane component 1 in menopause[J]. The Journal of Biomedical Research, 2021, 35(3): 228-237. DOI: 10.7555/JBR.35.20200172
Citation:
Lee Sang R., Yang Hyun, Jo Seong Lae, Lee Young Ho, Lee Hye Won, Park Bae-keun, Hong Eui-Ju. Suppressed estrogen supply via extra-ovarian progesterone receptor membrane component 1 in menopause[J]. The Journal of Biomedical Research, 2021, 35(3): 228-237. DOI: 10.7555/JBR.35.20200172
Lee Sang R., Yang Hyun, Jo Seong Lae, Lee Young Ho, Lee Hye Won, Park Bae-keun, Hong Eui-Ju. Suppressed estrogen supply via extra-ovarian progesterone receptor membrane component 1 in menopause[J]. The Journal of Biomedical Research, 2021, 35(3): 228-237. DOI: 10.7555/JBR.35.20200172
Citation:
Lee Sang R., Yang Hyun, Jo Seong Lae, Lee Young Ho, Lee Hye Won, Park Bae-keun, Hong Eui-Ju. Suppressed estrogen supply via extra-ovarian progesterone receptor membrane component 1 in menopause[J]. The Journal of Biomedical Research, 2021, 35(3): 228-237. DOI: 10.7555/JBR.35.20200172
Eui-Ju Hong, College of Veterinary Medicine, 99 Daehak-ro, Suite 401Veterinary medicine Bldg., Yuseong, Daejeon 34134, Republic of Korea. Tel/Fax: +82-42-821-6781/+82-42-821-8903, Email: ejhong@cnu.ac.kr
In post-menopausal women, intra-mammary estrogen, which is converted from extra-ovarian estrone (E1), promotes the growth of breast cancer. Since the aromatase inhibitor letrozole does not suppress 17β-estradiol (E2) production from E1, high intra-mammary E1 concentrations impair letrozole's therapeutic efficacy. Progesterone receptor membrane component 1 (Pgrmc1) is a non-classical progesterone receptor associated with breast cancer progression. In the present study, we introduced a Pgrmc1 heterozygous knockout (hetero KO) murine model exhibiting low Pgrmc1 expression, and observed estrogen levels and steroidogenic gene expression. Naïve Pgrmc1 hetero KO mice exhibited low estrogen (E2 and E1) levels and low progesterone receptor (PR) expression, compared to wild-type mice. In contrast, Pgrmc1 hetero KO mice that have been ovariectomized (OVX), including letrozole-treated OVX mice (OVX-letrozole), exhibited high estrogen levels and PR expression. Increased extra-ovarian estrogen production in Pgrmc1 hetero KO mice was observed with the induction of steroid sulfatase (STS). In MCF-7 cell, letrozole suppressed PR expression, but PGRMC1 knockdown increased PR and STS expression. Our presented results highlight the important role of Pgrmc1 in modulating estrogen production when ovary-derived estrogen is limited, thereby suggesting a potential therapeutic approach for letrozole resistance.
The female sex hormone 17β-estradiol (E2) plays a significant role in the menstrual cycle. It is synthesized by ovarian granulosa cells via aromatization of androgen produced by ovarian theca cells[1]. A series of enzymes, comprising P450 and hydroxysteroid dehydrogenases (HSD) family proteins, are responsible for converting cholesterol to E2[2]. However, after menopause, E2 levels decrease significantly[3]. Accordingly, estrone (E1) represents the majority of estrogen[4] in the post-menopausal stage, due to synthesis from androstenedione and estrone sulfates (E1S) in the adipose tissue[5–6]. Locally-produced E1 is then converted to E2 by 17β-hydroxysteroid dehydrogenase enzymes[7]. Circulating extra-ovarian steroid hormones produced after cessation of ovarian function exist largely as sulfate-bound molecules. Sulfate-bound hormones are biologically inactive, but can be activated via steroid sulfatase (STS) mediated hydrolysis[8]. Local estrogen production can also be regulated by aromatase, which converts peripherally-derived androgen to estrogen. Aromatase may be pharmacologically inhibited using the anti-cancer drug letrozole[9].
Progesterone receptor membrane component 1 (Pgrmc1) differs from the classical progesterone receptor (PR) in that it localizes not to the nucleus but to the plasma membrane and endoplasmic reticulum. Pgrmc1 has unique structure which shows haem-dependent dimerization[10]. Unlike transcriptional activity of the nuclear PR, Pgrmc1 instead has metabolic relevance, including associations with the insulin receptor and phosphoenolpyruvate carboxylase expression[11–12]. In addition, Pgrmc1 regulates cholesterol synthesis by binding cytochrome P450 enzymes[13]. However, its impact on hormone synthesis and turnover remains speculative[14]. In a recent study, Pgrmc1 stimulated breast cancer cell growth by increasing estrogen synthesis[15], which is consistent with a previous result suggesting that Pgrmc1 enhances estrogen-induced proliferation[16]. Although the relationship between mammary tumors and Pgrmc1 is likely mediated by estrogen, the significance of Pgrmc1 function in mammary glands after cessation of ovarian function has not been discussed.
The present study introduces a female Pgrmc1 heterozygous knockout (hetero KO) murine model exhibiting low Pgrmc1 expression. Estrogen levels and steroidogenic gene expression were observed in order to focus on the steroid regulatory role of Pgrmc1 in female reproductive tissues other than the ovaries, especially when estrogens derive only from local sources. To eliminate ovarian estrogen, Pgrmc1 hetero KO mice were ovariectomized (OVX mice) and received letrozole pellet subcutaneous implants two weeks later (OVX-letrozole mice). The mice were sacrificed after ten weeks of letrozole administration. Compared to wild-type (WT) mice, Pgrmc1 hetero KO mice exhibited low estrogen levels. However, OVX and OVX-letrozole mice exhibited estrogen levels higher than those of WT mice, suggesting that low expression of Pgrmc1 encourages extra-ovarian estrogen production. This effect may be especially important under conditions requiring therapeutic letrozole usage.
Materials and methods
Animals
Female mice on a C57BL/6J background were housed in a pathogen-free facility at Chungnam National University under a standard 12-hour light:12-hour dark cycle, and fed standard chow with water provided ad libitum. Pgrmc1 hetero KO mice were obtained from our previously established line[17]. All mouse experiments were approved and performed under the Chungnam Facility Animal Care Committee (202006A-CNU-105). The mice were sacrificed by CO2 asphyxiation. For long-term OVX and letrozole injection, the mice were bilaterally ovariectomized for 2 weeks and subsequently inserted with letrozole pellet [Innovative Research of America, USA; 0.1 mg/(kg·day)] as previously described[18]. The mice were sacrificed after 10 weeks of letrozole pellet insert. The number of mice used for long-term experiment is 3 for each group. For short-term OVX, the mice were bilaterally ovariectomized and sacrificed after 2 weeks. For short-term letrozole injection, the mice were injected with letrozole dissolved in 10% dimethyl sulfoxide (intraperitoneal, 10 µg/day for 3 days) and sacrificed after 3 days. The number of mice used for short-term experiment is 3, 3, 4, 4, 3, and 3 (for respective groups; naïve WT, naïve Pgrmc1 hetero KO, letrozole-treated WT, letrozole-treated Pgrmc1 hetero KO, OVX WT, and OVX Pgrmc1 hetero KO).
RNA isolation and quantitative reversetranscription PCR
RNA was extracted from tissues and MCF7 cells by using TRIzol Reagent (Thermo Fisher Scientific, USA), chloroform (Sigma, USA), isopropanol (Merck, Germany), and DEPC (Amresco, USA). Following the manufacturer's protocol, cDNA was synthesized with 1 µg of total RNA and Reverse transcriptase kit (SG-cDNAS100, Smartgene, UK). Quantitative PCR was carried out using specific primers (Table 1), Excel Taq Q-PCR Master Mix (SG-SYBR-500, Smartgene), and Stratagene Mx3000P (Agilent Technologies) equipped with a 96-well optical reaction plate. All experiments were repeated in triplicate, and mRNA values were calculated based on the cycle threshold and monitored for a melting curve.
Table
1.
Primers used for quantitative reverse transcription PCR analysis
All cell culture reagents were purchased from Welgene (Gyungsan, Korea). MCF7 human breast cancer cells were maintained at 37 °C in a 5% CO2 atmosphere in DMEM (Welgene) supplemented with 5% (vol/vol) fetal bovine serum, penicillin (100 U/mol), and streptomycin (100 μg/mL). For PGRMC1 knockdown, siRNA transfection was performed using the Lipofectamine 2000 reagent (Thermo Fisher Scientific) according to the manufacturer's protocol. Negative control siRNA and PGRMC1 siRNA #1 and #2 were purchased from Bioneer (Daejeon, Korea). The sense sequences of PGRMC1 siRNA #1 and #2 were 5′-CAGUACAGUCGCUAGUCAA-3′ and 5′-CAGUUCACUUUCAAGUAUCAU-3′.
Western blotting
Protein was extracted from tissues and MCF7 cells by homogenization. Protein was proceeded to SDS-PAGE. Gels were blotted to PVDF membrane, and the membrane was blocked and incubated with primary antibodies: Rabbit polyclonal antibodies to β-actin (Santa Cruz, USA), PR (Santa Cruz), and STS (Proteintech, USA); Rabbit monoclonal antibody to PGRMC1 (CST, USA). After overnight incubation, the membranes were washed and incubated with secondary antibodies (anti-rabbit, Jackson laboratory, USA). Bands were observed with ECL solution (Cyanagen, Italy) after 3 times of wash.
Immunofluorescence
Slides were obtained by 4 to 5 µm section of the paraffin block and incubated in xylene for overnight. The slides were then processed to following hydration steps, including 100% to 70% ethanol and distilled water. Antigen retrieval was performed with 0.1% sodium citrate buffer (Georgiachem, USA) at 95 ˚C for 60 minutes. After cooling down, the slides were washed once with TBS-T and blocked with 3% bovine serum albumin. Primary antibodies (PR and STS) were incubated overnight at 4 °C. The slides were washed with TBS-T for 3 times and incubated with anti-rabbit secondary antibodies (Life technologies, USA) for 4 hours, room temperature.
E2 and E1 measurements
Plasma E2 and E1 were measured by E2 ELISA kit (ADI-900-174, Enzo Life Sciences) and E1 ELISA kit (Abnova, China) following manufacturer's protocol.
Statistical analysis
Data are reported as mean±SD. Student's t-test obtained differences between means, and the one-way ANOVA followed by a Tukey's multiple comparison test was performed using Graph Pad Software (GraphPad Inc., USA).
Results
Low levels of Pgrmc1 decreased ovarian estrogen synthesis
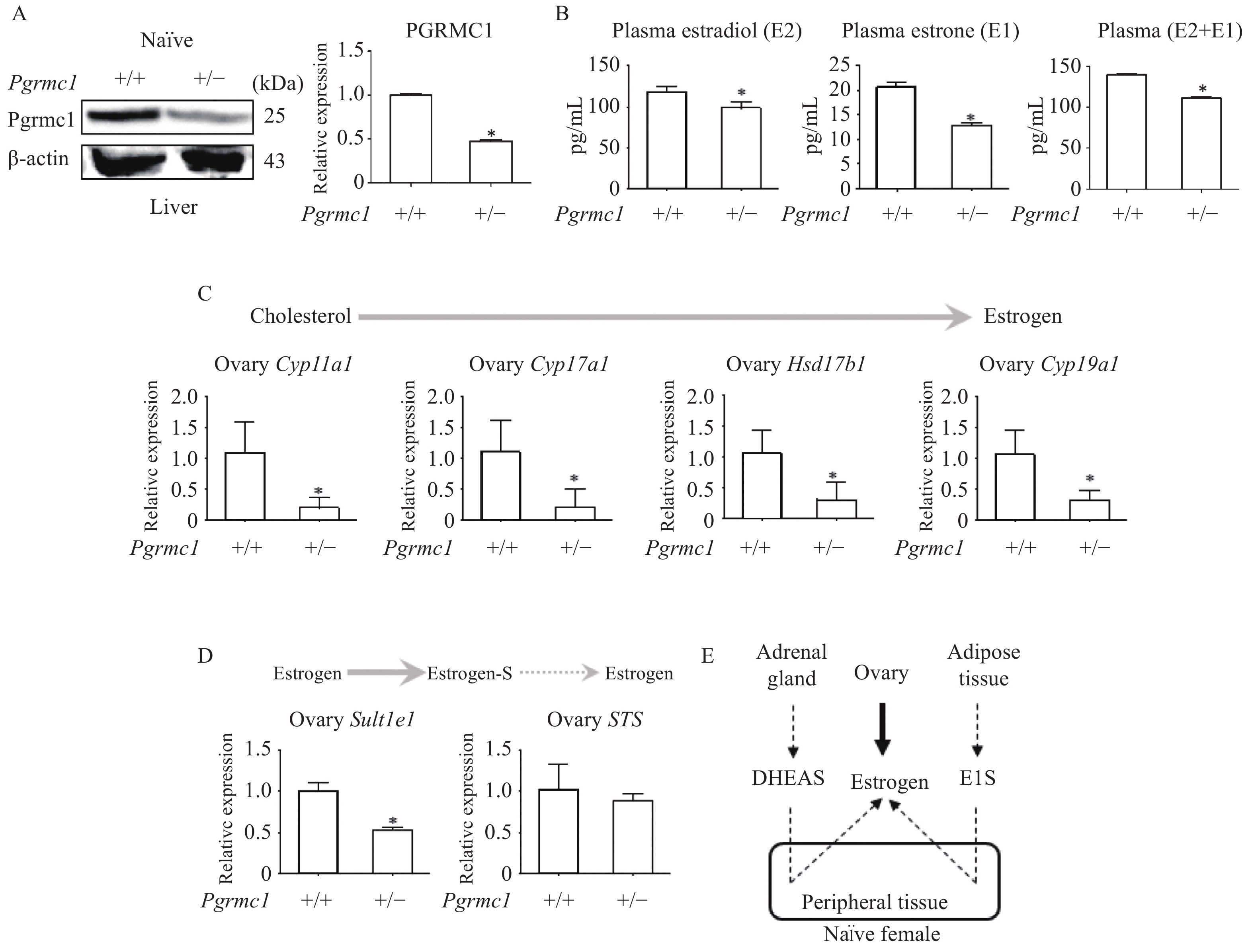
Adult female WT and Pgrmc1 hetero KO mice housed together (i.e. on a similar estrous cycle) were sacrificed, and hepatic Pgrmc1 expression levels were measured. Hepatic Pgrmc1 expression was significantly lower in Pgrmc1 hetero KO mice (47.6% of WT expression, P<0.05) (Fig. 1A). Regarding estrogen profiling (Fig. 1B), plasma E2 levels of Pgrmc1 hetero KO mice were significantly lower (82.9% of WT levels, P<0.05). Similarly, plasma E1 levels were also lower (61.6% of WT levels, P<0.05) (Fig. 1B). When combining E2 and E1, total non-pregnant estrogen levels were also lower (79.7% of WT levels, P<0.05) (Fig. 1B).
Figure
1.
Naïve Pgrmc1 heterozygous knockout mice produced less ovarian estrogen.
A: Western blotting analysis and quantification of Pgrmc1 in the livers of naïve wild-type (WT) and Pgrmc1 heterozygous knockout (hetero KO) mice. β-actin was used for an internal control. B: Plasma 17β-estradiol (E2) and estrone (E1) levels in naïve WT and Pgrmc1 hetero KO mice. C: mRNA expression of estrogen synthesis genes in ovaries of naïve WT and Pgrmc1 hetero KO mice. D: mRNA expression of Sult1e1 and STS in ovaries of naïve WT and Pgrmc1 hetero KO mice. E: Illustrated pathway of estrogen synthesis in naïve female mice. Values are reported as mean±SD. Student's t-test was performed to indicate significance. *P<0.05 vs. naïve WT (n=3). Solid line indicates major metabolism while dashed line indicates minor metabolism. DHEAS: dehydroepiandrosterone sulfate.
In adult female mice with normal ovarian function, the majority of estrogen is synthesized within the ovary. Regarding estrogen biosynthetic pathway enzymes, levels of ovarian transcripts, including Cyp11a1, Cyp17a1, Hsd17b1, and Cyp19a1, were significantly lower in Pgrmc1 hetero KO mice (17%, 17.4%, 28.2%, and 28.7%, respectively of WT levels; P<0.05) (Fig. 1C). Consistent with these findings, levels of ovarian estrogen sulfation gene (Sult1e1) transcripts were also lower (52.7% of WT levels, P<0.05) (Fig. 1D). Meanwhile, ovarian transcription of STS, which responsible for estrogen sulfate hydrolysis, did not differ significantly between Pgrmc1 hetero KO and WT mice (Fig. 1D). The normal murine estrogen biosynthetic pathway is illustrated in Fig. 1E.
Low levels of Pgrmc1 increased extra-ovarian estrogen synthesis in OVX and OVX letrozole-treated mice
Lack of ovaries represents a specific condition under which extra-ovarian estrogen synthesis becomes pronounced. The OVX experimental schedule is shown in Fig. 2A. Western blotting analysis demonstrated that hepatic Pgrmc1 expression was significantly lower in OVX Pgrmc1 hetero KO mice (42.1% of OVX WT mouse levels, P<0.05) (Fig. 2B). Relative to normal WT mice, in which E2 levels varied within the range 110 to 120 pg/mL, E2 levels were much lower in OVX WT mice, which exhibited a range of 70 to 80 pg/mL. Relative to OVX WT mice, E2 levels significantly increased in OVX Pgrmc1 hetero KO mice (1.12-fold higher than those of OVX WT mice, P<0.05) (Fig. 2C). Furthermore, the increase in plasma E1 level in OVX Pgrmc1 hetero KO mice was marked (1.58-fold higher than that of OVX WT mice, P<0.05) (Fig. 2C). Accordingly, total plasma estrogen level also increased in OVX Pgrmc1 hetero KO mice (1.28-fold higher than that of OVX WT mice, P<0.05) (Fig. 2C). The estrogen biosynthetic pathway in ovariectomized mice is illustrated in Fig. 2D.
Figure
2.
OVX and OVX-letrozole Pgrmc1 heterozygous knockout mice produced more extra-ovarian estrogen.
A and E: Experimental schedule for long-term OVX and letrozole pellet administration in wild-type (WT) and Pgrmc1 heterozygous knockout (hetero KO) mice. B and F: Western blotting analysis and quantification of Pgrmc1 in the livers of OVX and OVX-letrozole WT and Pgrmc1 hetero KO mice. β-actin was used for an internal control. C and G: Plasma 17β-estradiol (E2) and estrone (E1) levels in OVX and OVX-letrozole WT and Pgrmc1 hetero KO mice. D and H: Illustrated pathway of estrogen synthesis in OVX and OVX-letrozole female mice. Values are reported as mean±SD. Student's t-test was performed to indicate significance. *P<0.05 vs. OVX WT (n=3) or OVX-letrozole WT (n=3). OVX: ovariectomized; DHEAS: dehydroepiandrosterone sulfate.
To exclude the aromatase regulatory mechanism, an aromatase inhibitor, letrozole, was subcutaneously embedded in pellet-form. Because of the slow-release effect of letrozole pellets, mice were sacrificed ten weeks after letrozole implantation. The experimental schedule is described in Fig. 2E. Hepatic Pgrmc1 expression of OVX-letrozole Pgrmc1 hetero KO mice remained significantly lower (91.9% that of OVX-letrozole WT mice, P<0.05) (Fig. 2F), although the difference is far less pronounced than when comparing OVX Pgrmc1 hetero KO to OVX WT mice. Nevertheless, plasma E2 levels were higher in OVX-letrozole Pgrmc1 hetero KO mice (1.17-fold higher than those of OVX-letrozole WT mice, P<0.05) (Fig. 2G). While E1 levels were similar, total estrogen (E2+E1) levels thus remained were higher in OVX-letrozole Pgrmc1 hetero KO mice (1.17-fold higher than those of OVX-letrozole WT mice, P<0.05) (Fig. 2G). These findings suggest that Pgrmc1 suppresses local estrogen production regardless of aromatase activity. The estrogen biosynthetic pathway in OVX-letrozole mice is illustrated in Fig. 2H.
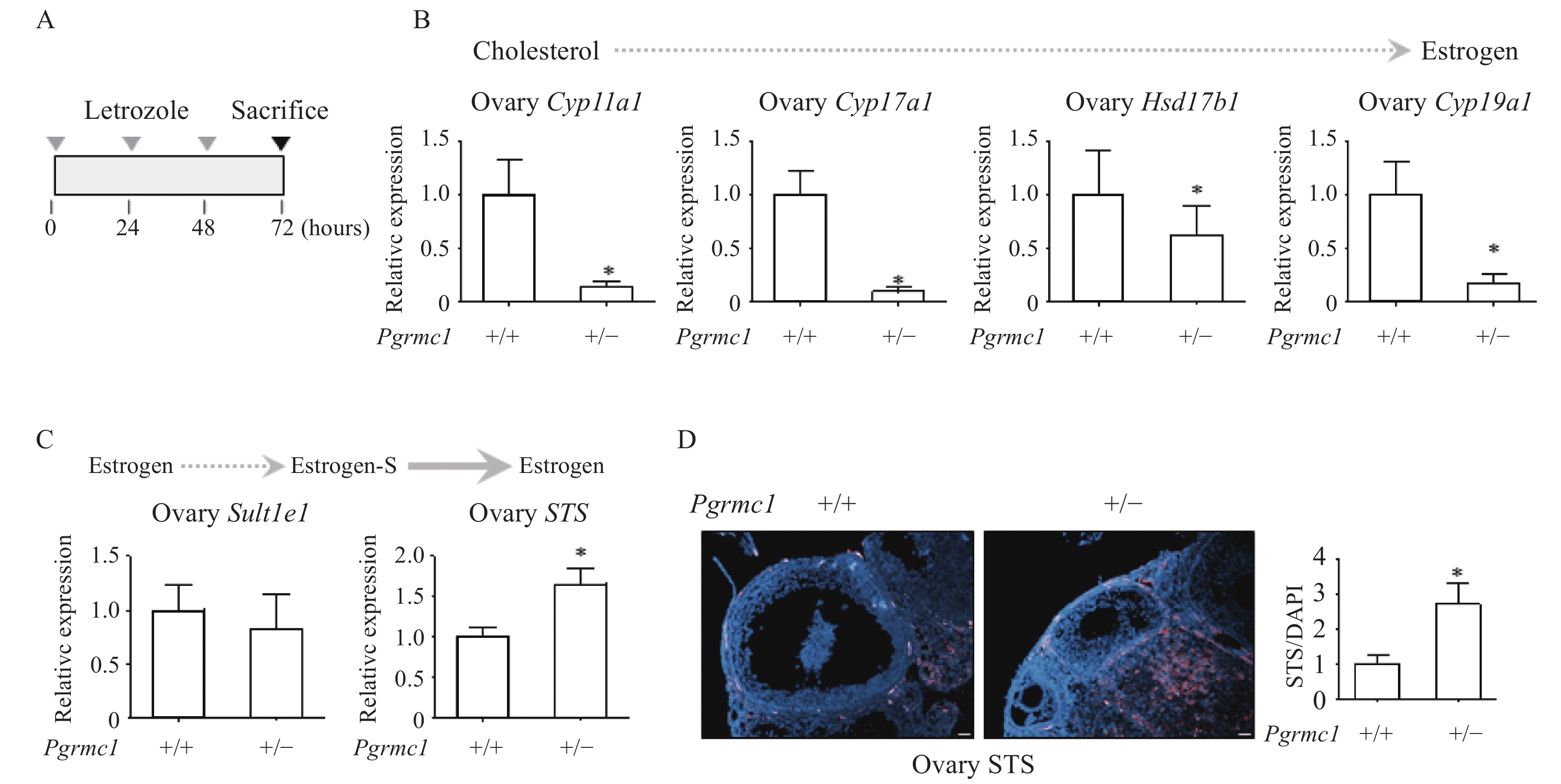
Low levels of Pgrmc1 increased ovarian STS expression in letrozole-treated mice
Given that OVX-letrozole mice lack ovaries, an additional letrozole-only group was analyzed for ovarian estrogen biosynthetic pathway gene expression. Short-term letrozole treatment (10 µg per day for three consecutive days) induced abrupt gene expression changes (Fig. 3A). Transcript levels of Cyp11a1, Cyp17a1, Hsd17b1, and Cyp19a1 were significantly lower in ovaries of Pgrmc1 hetero KO mice receiving letrozole (13.9%, 9.2%, 62.7%, and 17.0%, respectively, of those of letrozole-treated WT mice; P<0.05) (Fig. 3B). However, Sult1e1 transcript levels were similar, while those of STS were significantly higher in ovaries of Pgrmc1 hetero KO mice receiving letrozole (1.65-fold higher than those of letrozole-treated WT mice, P<0.05) (Fig. 3C). Furthermore, immunostaining demonstrated that STS expression was significantly higher in ovaries of letrozole-treated Pgrmc1 hetero KO mice (2.7-fold higher than that of letrozole-treated WT mice, P<0.05) (Fig. 3D).
A: Experimental schedule for short-term letrozole injection in wild-type (WT) and Pgrmc1 heterozygous knockout (hetero KO) mice. B: mRNA expression of estrogen synthesis genes in ovaries of letrozole-treated WT and Pgrmc1 hetero KO mice. C: mRNA expression of Sult1e1 and STS in ovaries of letrozole-treated WT and Pgrmc1 hetero KO mice. D: Immunostaining analysis and quantification of STS in the ovaries of letrozole-treated WT and Pgrmc1 hetero KO mice (scale bar=400 μm). STS (pink) positive signals were normalized to DAPI (blue). Image J was used for quantification. Values are reported as mean±SD. Student's t-test was performed to indicate significance. *P<0.05 vs. letrozole-treated WT (n=4). Solid line indicates major metabolism while dashed line indicate minor metabolism.
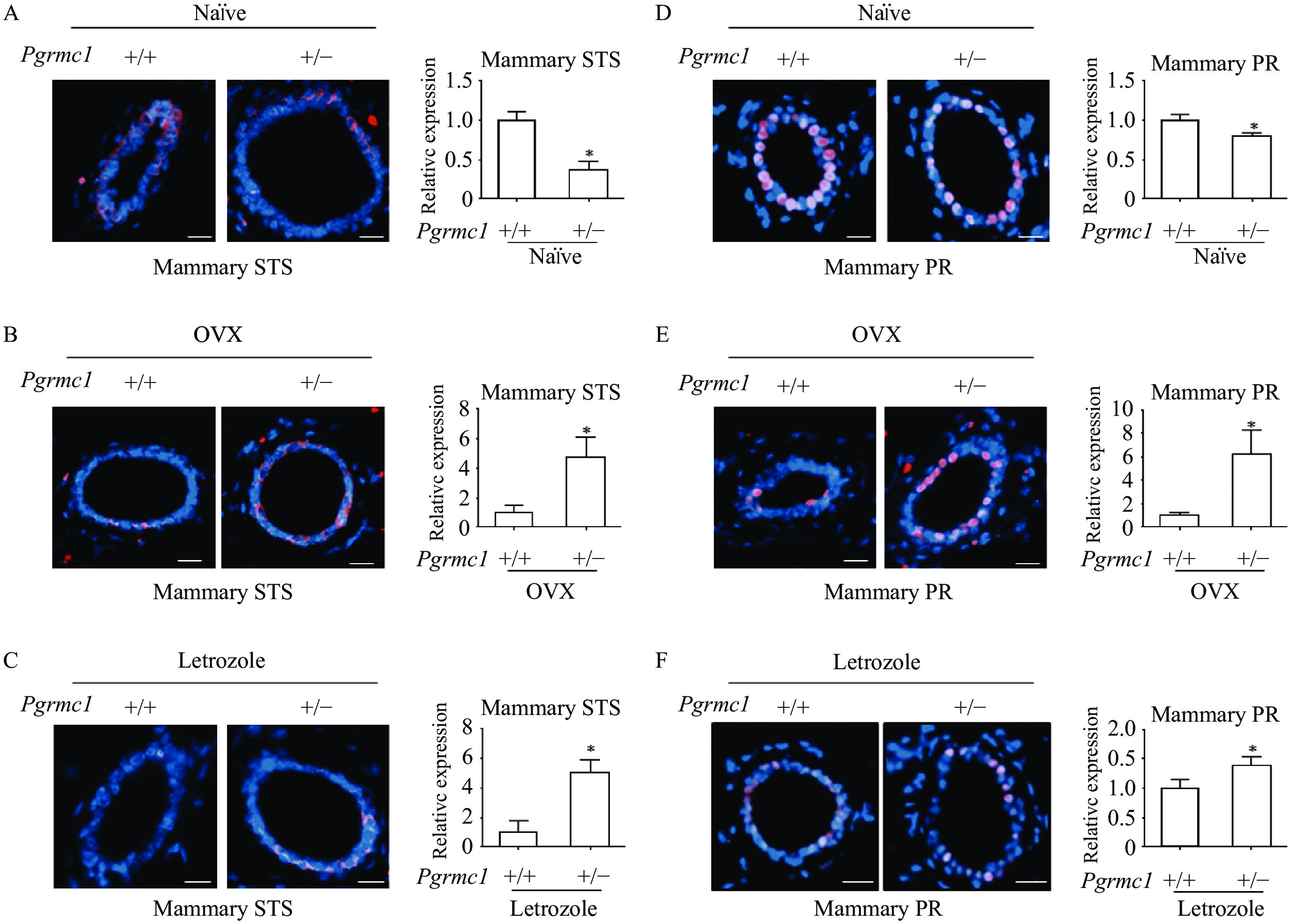
Low levels of Pgrmc1 increased mammary gland estrogenic capacity in OVX and letrozole-treated mice via enhanced STS expression
Given that ovaries of letrozole-treated mice exhibited high STS expression, we sought to investigate whether this enzyme is also involved in local estrogen production in the mammary gland in Pgrmc1 hetero KO mice. Due to the difficulty in cleanly dissecting mammary tissue, STS expression was measured by immunostaining. Mammary gland expression of STS in Pgrmc1 hetero KO mice was significantly lower (37% that of WT mice, P<0.05) (Fig. 4A). Conversely, mammary gland expression of STS in OVX and letrozole-treated Pgrmc1 hetero KO mice was significantly higher (4.73- and 5.02-fold higher, respectively, than that of OVX and letrozole-treated WT mice, P<0.05) (Fig. 4B–C). As a marker of estrogen activity, mammary gland expression of PR in Pgrmc1 hetero KO mice was lower (80.3% that of WT mice, P<0.05) (Fig. 4D). Conversely, mammary gland expression of PR in OVX and letrozole-treated Pgrmc1 hetero KO mice was higher (6.25- and 1.4-fold higher, respectively, that than of OVX and letrozole-treated WT mice, P<0.05) (Fig. 4E–F). Findings regarding STS expression suggest that conversion of estrogen sulfate supplements local estrogen production in the mammary glands of OVX and letrozole-treated Pgrmc1 hetero KO mice.
A and D: Immunostaining analysis and quantification of STS and PR in the mammary glands of naïve wild-type (WT) and Pgrmc1 heterozygous knockout (hetero KO) mice (scale bar=200 μm). B and E: Immunostaining analysis and quantification of STS and PR in the mammary glands of OVX WT and Pgrmc1 hetero KO mice (scale bar=200 μm). C and F: Immunostaining analysis and quantification of STS and PR in the mammary glands of letrozole-treated WT and Pgrmc1 hetero KO mice (scale bar=200 μm). STS and PR (pink) positive signals were normalized to DAPI (blue). Image J was used for quantification. Values are reported as mean±SD. Student's t-test was performed to indicate significance. *P<0.05 vs. naïve WT (n=3) or OVX WT (n=3) or letrozole-treated WT (n=4). STS: steroid sulfatase; PR: progesterone receptor.
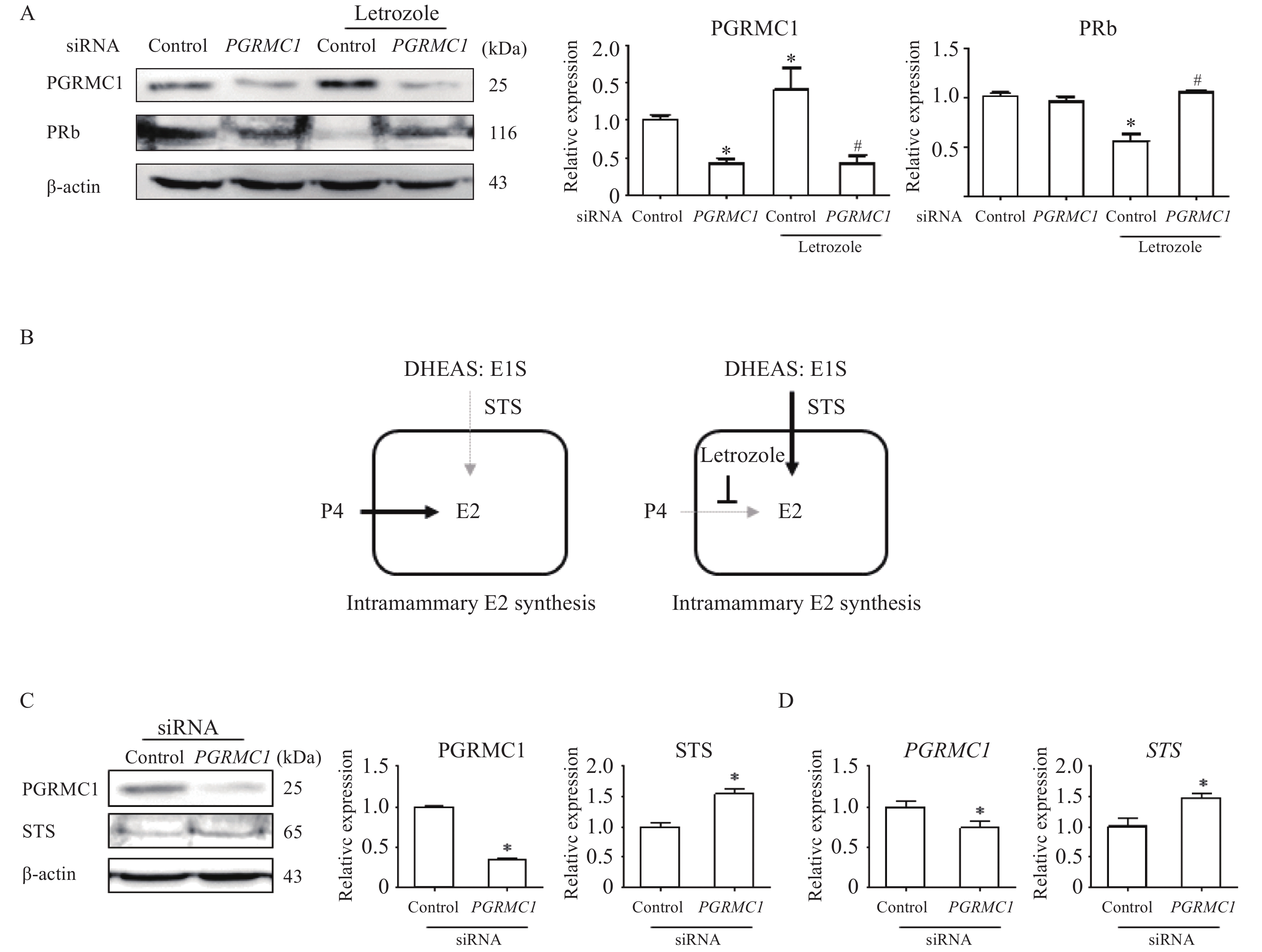
Low levels of PGRMC1 increased estrogenic capacity in MCF7 cells, thereby inducing letrozole resistance
To evaluate the role of Pgrmc1 in modulating endocrine factors other than estrogen level, an in vitro experiment was conducted using the MCF7 cell line in which estrogen-estrogen receptor signaling is vigorous. Expression of PGRMC1 was knocked down using siRNA. During cell culture, progesterone pre-treatment (10 nmol/L for 24 hours) to provide estrogen and estrogen sulfate precursors occurred prior to letrozole treatment (100 nmol/L for 24 hours). Cells were then harvested for analysis. Knockdown-mediated suppression of PGRMC1 expression was confirmed (42.7% of control cell levels, P<0.05) (Fig. 5A). When letrozole treatment increased PGRMC1 expression in control cells (1.41-fold higher than vehicle-treated control cells, P<0.05), the letrozole-treated knockdown group instead exhibited decreased expression (30.2% that of letrozole-treated control cells, P<0.05) (Fig. 5A). As a marker of estrogen activity, PRb expression in letrozole-treated WT cells was significantly lower (55.9% that of vehicle-treated WT cells, P<0.05) (Fig. 5A). However, in the knockdown group, letrozole treatment instead increased PRb expression (1.88-fold higher than that of letrozole-treated control cells, P<0.05), while letrozole treatment did not alter PR expression within the knockdown group itself (Fig. 5A). These results may be attributed to STS impact on steroid hormone metabolism (Fig. 5B). Indeed, when PGRMC1 expression was suppressed via knockdown (35.3% that of control cells, P<0.05), expression of STS increased (1.54-fold higher than that of control cells, P<0.05) (Fig. 5C). Concomitantly, knockdown decreased PGRMC1 transcription (74% that of control cells, P<0.05) and increased STS transcription (1.48-fold higher than that of control cells, P<0.05) (Fig. 5D).
Figure
5.PGRMC1 suppression increased PR and STS expression in MCF7 cells.
A: Western blotting analysis and quantification of PGRMC1 and PRb in vehicle or letrozole-treated control and PGRMC1 siRNA groups. β-actin was used for an internal control. B: Illustrated pathway for estrogen production in letrozole-treated MCF7 cells. C: Western blotting analysis and quantification of PGRMC1 and STS in control and PGRMC1 siRNA groups. β-actin was used for an internal control. D: mRNA expression of PGRMC1 and STS in control and PGRMC1 siRNA groups. RPLP0 was used for internal control. Values are reported as means±SD. One-way ANOVA followed by a Tukey's multiple comparison test (A) or Student's t-test (C and D) was performed to indicate significance. *P<0.05 vs. control siRNA group. #P<0.05 vs. letrozole-treated control siRNA group. In vitro experiments were repeated at least 3 times. DHEAS: dehydroepiandrosterone sulfate; E1S: estrone sulfates; STS: steroid sulfatase; E2: 17β-estradiol.
Previous studies suggest that Pgrmc1 may play a critical role in mammary tumor growth mediated by estrogen ligation of ERα. In breast cancer patients, Pgrmc1 levels correlate with ERα expression[19]. Additionally, Pgrmc1 sensitizes estrogen-induced proliferation of MCF7 cells[20] and induces mammary tumor growth in a xenograft model via its estrogenic effect[15]. Nonetheless, it is not known whether Pgrmc1 modulates mammary tumor growth when endogenous estrogen supply is limited, such as post-menopausally, after OVX, or in response to letrozole treatment. The present study focused on the role of Pgrmc1 when ovarian estrogen is eliminated via surgery (OVX) or when levels of estrogen are decreased via letrozole-mediated aromatase inhibition. Results demonstrate that Pgrmc1 suppresses plasma estrogen levels and intra-mammary estrogen levels via suppressed STS expression.
Letrozole is an anti-cancer drug indicated for hormone-sensitive breast cancer in post-menopausal women. Its therapeutic mechanism is based on highly-selective inhibition of aromatase, without impacting other steroidogenic enzymes. Inhibition of aromatization consequently decreases estrogen levels, but certain tumors exhibit letrozole resistance. It has previously been demonstrated that letrozole resistance depends on expression of estrogen-regulated and proliferative genes[21]. Moreover, sensitivity and responses to letrozole are dependent on estrogen and progesterone receptor status[22]. Accordingly, both estrogen receptor dysfunction and the presence of alternative estrogen sources can lead to letrozole resistance[23–24]. Compared to WT mice, Pgrmc1 hetero KO mice demonstrated low levels of ovarian estrogen synthesis. However, when Pgrmc1 hetero KO mice underwent OVX and letrozole treatment, estrogen levels unexpectedly increased relative to WT mice. Importantly, letrozole treatment of Pgrmc1 hetero KO mice increased mammary gland PR expression, thereby increasing estrogenic capacity. Consistent with these observations, MCF7 cells which had undergone Pgrmc1 knockdown exhibited an increase in PR expression in response to letrozole treatment. These results suggest that decreased Pgrmc1 expression increases estrogenesis even in the absence of ovarian estrogen synthesis and when androgen aromatization is limited. This may have important therapeutic implications in increasing breast cancer sensitivity to letrozole.
The mechanism by which low Pgrmc1 expression increases estrogenesis in OVX and letrozole-treated conditions is hypothesized. Post-menopausally, peripheral tissue aromatization increases, although plasma estrogen concentration remains low. Because estrogen derived from aromatized testosterone can trigger breast cancer, letrozole can effectively suppress breast cancer[23]. Simultaneously, peripheral tissues, (including the adrenal gland and adipose tissue) produce inactive estrogen and androgen forms post-menopausally. These less active forms of steroid hormones are mostly sulfate-bound. Interestingly, estrone sulfate can be converted to E1 and E2 in healthy breast parenchymal tissue[25]. Post-menopausal women produce estrogen, especially intra-mammary estrogen, largely from such sulfate-bound forms of steroid hormone[26]. The enzyme responsible for the hydrolysis of sulfate from E1S, thereby converting it to active E1, is STS[27]. Moreover, this estrogenic effect of STS has been demonstrated in post-menopausal breast cancer patients[28]. Therefore, targeting of STS has been discussed as a therapeutic strategy to inhibit the growth of estrogen-dependent breast cancers[29]. Since letrozole inhibits only aromatization of androgen to estrogen, it implicitly does not suppress estrogen production via the sulfatase pathway. Moreover, higher STS levels have been observed in aromatase-inhibited breast cancer patients[30]. For these reasons, many research groups have focused on the dual inhibition of aromatase and sulfatase to suppress breast cancer[8]. Although ovarian Pgrmc1 increases E2 synthesis from cholesterol, mammary Pgrmc1 suppresses STS expression when the cholesterol-E2 pathway is inhibited. Therefore, the present study suggests that Pgrmc1 is a novel therapeutic target in letrozole-treated patients.
Pgrmc1 has been suggested as a mammary tumor prognostic marker associated with estrogenic conditions[31]; in agreement, the present study demonstrated that Pgrmc1 is associated with estrogen synthesis in mice. Low estrogenic conditions in Pgrmc1 hetero KO mice explain results of a previous study in which Pgrmc1 KO suppressed mammary gland development[32]. Furthermore, the present study demonstrated that a low level of Pgrmc1 results in estrogen maintenance in OVX and letrozole-treated mice via STS induction. Therefore, the present study highlights the contradictory role of Pgrmc1 in estrogen regulation and suggests a novel therapeutic approach for ameliorating letrozole-resistance in post-menopausal breast cancer patients.
Acknowledgments
This work was supported by a research fund of Chungnam National University (No. 2020-0733-01). This work was supported by Research Scholarship of Chungnam National University.
Cui J, Shen Y, Li R. Estrogen synthesis and signaling pathways during aging: from periphery to brain[J]. Trends Mol Med, 2013, 19(3): 197–209. doi: 10.1016/j.molmed.2012.12.007
[3]
Khosla S, Atkinson EJ, Melton Ⅲ LJ, et al. Effects of age and estrogen status on serum parathyroid hormone levels and biochemical markers of bone turnover in women: a population-based study[J]. J Clin Endocrinol Metab, 1997, 82(5): 1522–1527. doi: 10.1210/jcem.82.5.3946
[4]
Vermeulen A, Verdonck L. Factors affecting sex hormone levels in postmenopausal women[J]. J Steroid Biochem, 1979, 11(1): 899–904. doi: 10.1016/0022-4731(79)90027-X
[5]
Forney JP, Milewich L, Chen GT, et al. Aromatization of androstenedione to estrone by human adipose tissue in vitro. Correlation with adipose tissue mass, age, and endometrial neoplasia[J]. J Clin Endocrinol Metab, 1981, 53(1): 192–199. doi: 10.1210/jcem-53-1-192
[6]
Hetemäki N, Savolainen-Peltonen H, Tikkanen MJ, et al. Estrogen metabolism in abdominal subcutaneous and visceral adipose tissue in postmenopausal women[J]. J Clin Endocrinol Metab, 2017, 102(12): 4588–4595. doi: 10.1210/jc.2017-01474
[7]
Marchais-Oberwinkler S, Henn C, Möller G, et al. 17β-Hydroxysteroid dehydrogenases (17β-HSDs) as therapeutic targets: protein structures, functions, and recent progress in inhibitor development[J]. J Steroid Biochem Mol Biol, 2011, 125(1–2): 66–82. doi: 10.1016/j.jsbmb.2010.12.013
[8]
Reed MJ, Purohit A, Woo LWL, et al. Steroid sulfatase: molecular biology, regulation, and inhibition[J]. Endocr Rev, 2005, 26(2): 171–202. doi: 10.1210/er.2004-0003
[9]
Lamb HM, Adkins JC. Letrozole: A review of its use in postmenopausal women with advanced breast cancer[J]. Drugs, 1998, 56(6): 1125–1140. doi: 10.2165/00003495-199856060-00020
[10]
Kabe Y, Nakane T, Koike I, et al. Haem-dependent dimerization of PGRMC1/Sigma-2 receptor facilitates cancer proliferation and chemoresistance[J]. Nat Commun, 2016, 7: 11030. doi: 10.1038/ncomms11030
[11]
Hampton KK, Anderson K, Frazier H, et al. Insulin receptor plasma membrane levels increased by the progesterone receptor membrane component 1[J]. Mol Pharmacol, 2018, 94(1): 665–673. doi: 10.1124/mol.117.110510
[12]
Lee SR, Choi WY, Heo JH, et al. Progesterone increases blood glucose via hepatic progesterone receptor membrane component 1 under limited or impaired action of insulin[J]. Sci Rep, 2020, 10(1): 16316. doi: 10.1038/s41598-020-73330-7
[13]
Hughes AL, Powell DW, Bard M, et al. Dap1/PGRMC1 binds and regulates cytochrome P450 enzymes[J]. Cell Metab, 2007, 5(2): 143–149. doi: 10.1016/j.cmet.2006.12.009
[14]
Rohe HJ, Ahmed IS, Twist KE, et al. PGRMC1 (progesterone receptor membrane component 1): a targetable protein with multiple functions in steroid signaling, P450 activation and drug binding[J]. Pharmacol Ther, 2009, 121(1): 14–19. doi: 10.1016/j.pharmthera.2008.09.006
[15]
Asperger H, Stamm N, Gierke B, et al. Progesterone receptor membrane component 1 regulates lipid homeostasis and drives oncogenic signaling resulting in breast cancer progression[J]. Breast Cancer Res, 2020, 22(1): 75. doi: 10.1186/s13058-020-01312-8
[16]
Li X, Ruan X, Gu M, et al. PGRMC1 can trigger estrogen-dependent proliferation of breast cancer cells: estradiol vs. equilin vs. ethinylestradiol[J]. Climacteric, 2019, 22(5): 483–488. doi: 10.1080/13697137.2019.1582624
[17]
Lee SR, Kwon SW, Kaya P, et al. Loss of progesterone receptor membrane component 1 promotes hepatic steatosis via the induced de novo lipogenesis[J]. Sci Rep, 2018, 8(1): 15711. doi: 10.1038/s41598-018-34148-6
[18]
Yang H, Lee SY, Lee SR, et al. Therapeutic effect of Ecklonia cava extract in letrozole-induced polycystic ovary syndrome rats[J]. Front Pharmacol, 2018, 9: 1325. doi: 10.3389/fphar.2018.01325
[19]
Zhang Y, Ruan XY, Willibald M, et al. May progesterone receptor membrane component 1 (PGRMC1) predict the risk of breast cancer?[J]. Gynecol Endocrinol, 2016, 32(1): 58–60. doi: 10.3109/09513590.2015.1078303
[20]
Neubauer H, Yang Y, Seeger H, et al. The presence of a membrane-bound progesterone receptor sensitizes the estradiol-induced effect on the proliferation of human breast cancer cells[J]. Menopause, 2011, 18(8): 845–850. doi: 10.1097/gme.0b013e31820e5ac5
[21]
Miller WR, Larionov A. Changes in expression of oestrogen regulated and proliferation genes with neoadjuvant treatment highlight heterogeneity of clinical resistance to the aromatase inhibitor, letrozole[J]. Breast Cancer Res, 2010, 12(4): R52. doi: 10.1186/bcr2611
[22]
Miller WR. Aromatase inhibitors: prediction of response and nature of resistance[J]. Expert Opin Pharmacother, 2010, 11(11): 1873–1887. doi: 10.1517/14656566.2010.487863
[23]
Bhatnagar AS. The discovery and mechanism of action of letrozole[J]. Breast Cancer Res Treat, 2007, 105(Suppl 1): 7–17. doi: 10.1007/s10549-007-9696-3
[24]
Elledge RM, Osborne CK. Oestrogen receptors and breast cancer[J]. BMJ, 1997, 314(7098): 1843–1844. doi: 10.1136/bmj.314.7098.1843
[25]
Chatterton Jr RT, Geiger AS, Gann PH, et al. Formation of estrone and estradiol from estrone sulfate by normal breast parenchymal tissue[J]. J Steroid Biochem Mol Biol, 2003, 86(2): 159–166. doi: 10.1016/S0960-0760(03)00266-8
[26]
Pasqualini JR, Gelly C, Nguyen BL, et al. Importance of estrogen sulfates in breast cancer[J]. J Steroid Biochem, 1989, 34(1–6): 155–163. doi: 10.1016/0022-4731(89)90077-0
[27]
Hanamura T, Niwa T, Gohno T, et al. Possible role of the aromatase-independent steroid metabolism pathways in hormone responsive primary breast cancers[J]. Breast Cancer Res Treat, 2014, 143(1): 69–80. doi: 10.1007/s10549-013-2788-3
[28]
Nakata T, Takashima S, Shiotsu Y, et al. Role of steroid sulfatase in local formation of estrogen in post-menopausal breast cancer patients[J]. J Steroid Biochem Mol Biol, 2003, 86(3–5): 455–460. doi: 10.1016/S0960-0760(03)00357-1
[29]
Geisler J, Sasano H, Chen S, et al. Steroid sulfatase inhibitors: promising new tools for breast cancer therapy?[J]. J Steroid Biochem Mol Biol, 2011, 125(1–2): 39–45. doi: 10.1016/j.jsbmb.2011.02.002
[30]
Chanplakorn N, Chanplakorn P, Suzuki T, et al. Increased estrogen sulfatase (STS) and 17β-hydroxysteroid dehydrogenase type 1(17β-HSD1) following neoadjuvant aromatase inhibitor therapy in breast cancer patients[J]. Breast Cancer Res Treat, 2010, 120(3): 639–648. doi: 10.1007/s10549-010-0785-3
[31]
Ruan XY, Cai GJ, Wei Y, et al. Association of circulating Progesterone Receptor Membrane Component-1 (PGRMC1) with breast tumor characteristics and comparison with known tumor markers[J]. Menopause, 2020, 27(2): 183–193. doi: 10.1097/GME.0000000000001436
[32]
Kim G, Lee JG, Cheong SA, et al. Progesterone receptor membrane component 1 is required for mammary gland development[J]. Biol Reprod, 2020, 103(6): 1249–1259. doi: 10.1093/biolre/ioaa164
Minbo J, Feng C, Wen H, et al. Up-regulated and hypomethylated genes are causative factors and diagnostic markers of osteoporosis. Am J Transl Res, 2023, 15(10): 6042-6057.
2.
Zhou Z, Huang Z, Khan HM, et al. Identification of 12 hub genes associated to the pathogenesis of osteoporosis based on microarray and single-cell RNA sequencing data. Funct Integr Genomics, 2023, 23(2): 186.
DOI:10.1007/s10142-023-01116-x
3.
Hassanshahi F, Noroozi Karimabad M, Miranzadeh E, et al. The Serum Level of CXCL9, CXCL10, and CXCL11 and the Expression of CXCR3 of Peripheral Blood Mononuclear Cells in Brucellosis Patients. Curr Microbiol, 2023, 80(6): 201.
DOI:10.1007/s00284-023-03230-2
4.
Karimabad MN, Hassanshahi G, Kounis NG, et al. The Chemokines CXC, CC and C in the Pathogenesis of COVID-19 Disease and as Surrogates of Vaccine-Induced Innate and Adaptive Protective Responses. Vaccines (Basel), 2022, 10(8): 1299.
DOI:10.3390/vaccines10081299
5.
Yuan SG, Hu HL, Wang XJ, et al. Bindarit Reduces Bone Loss in Ovariectomized Mice by Inhibiting CCL2 and CCL7 Expression via the NF-κB Signaling Pathway. Orthop Surg, 2022, 14(6): 1203-1216.
DOI:10.1111/os.13252
6.
Karimabad MN, Kounis NG, Hassanshahi G, et al. The Involvement of CXC Motif Chemokine Ligand 10 (CXCL10) and Its Related Chemokines in the Pathogenesis of Coronary Artery Disease and in the COVID-19 Vaccination: A Narrative Review. Vaccines (Basel), 2021, 9(11): 1224.
DOI:10.3390/vaccines9111224
7.
Zhu S, Tian A, Guo L, et al. Investigation of Diagnostic Biomarkers for Osteoporosis Based on Differentially Expressed Gene Profile with QCT and mDixon-Quant Techniques. Orthop Surg, 2021, 13(7): 2137-2144.
DOI:10.1111/os.13094
8.
Ahmadi H, Khorramdelazad H, Hassanshahi G, et al. Involvement of Eotaxins (CCL11, CCL24, CCL26) in Pathogenesis of Osteopenia and Osteoporosis. Iran J Public Health, 2020, 49(9): 1769-1775.
DOI:10.18502/ijph.v49i9.4098
9.
Janas A, Kruczek E, Londzin P, et al. Negligible Effect of Estrogen Deficiency on Development of Skeletal Changes Induced by Type 1 Diabetes in Experimental Rat Models. Mediators Inflamm, 2020, 2020: 2793804.
DOI:10.1155/2020/2793804
10.
Hosseini FS, Noroozi Karimabad M, Hajizadeh MR, et al. Evaluating of Induction of Apoptosis by Cornus mass L. Extract in the Gastric Carcinoma Cell Line (AGS). Asian Pac J Cancer Prev, 2019, 20(1): 123-130.
DOI:10.31557/APJCP.2019.20.1.123
 Authors and Reviewers
Authors and Reviewers

 DownLoad:
DownLoad: